


















文献精读 | DOCK7协同TACC3调节放射状胶质细胞区间动态核迁移及皮质发育




本文由“山中麻薯”授权转载

在神经发育过程中,神经前体细胞通过细胞增殖和分化形成神经元,进而形成皮层的六层结构。锥体神经元大部分由放射状神经胶质细胞(RGC)分化而来,而后者是高度极化的细胞,以区间动态核迁移(INM)为特征,即细胞核随细胞周期改变而沿顶-基轴规律移动。在细胞周期的G1期,细胞核从室面向顶端移动,在S期位于皮层基底端,在G2期返回室面,并在此处进行细胞分裂。
控制INM的分子机制涉及肌动蛋白和微管系统,前者通常与细胞核从室面向顶端迁移有关,而后者通常与细胞核从基底端向室面迁移有关。大量证据表明,中心体在细胞核从基底端向顶端迁移过程中至关重要,它组织了微管的形成。Cep120和TACC是微管中的两种蛋白,它们在微管中参与连接中心体和细胞核,抑制这两种蛋白的表达,可扰乱INM,并导致细胞分化减少增殖增加。尽管中心体蛋白在INM和维持细胞池中非常重要,但其具体机制尚不清楚。
1、DOCK7在发育皮层中的表达与位置
皮层western blot结果表明,DOCK7在胚胎第11天开始表达,并且持续到至少出生后第3天,这表明DOCK7在皮质神经发生的活跃期表达(图1a)。免疫组化结果显示E13.5时Tuj1+的神经元及Nestin+的VZ区前体细胞中高表达DOCK7(图1b),提示DOCK7表达的与神经发生密切相关。
值得注意的是,DOCK7在VZ区高表达于室管膜面,该部位Nestin阳性的前体细胞中,DOCK7与中心体标志物γ-tubulin共标,表明DOCK7存在于中心体(图1c),体外研究也证实了这一点(图1d)。这些数据证实了在皮质前体细胞中心体中表达DOCK7。

图1
2、出生前抑制DOCK7表达促进VZ区前体细胞增殖
Dock7#2或scr#1 shRNA(对照组)构建体以及EGFP-NLS质粒一起通过胚胎电转导入E13.5小鼠大脑,并在E15天进行BrdU染色,注射2小时后进行免疫荧光分析(图2)。Dock7#2 shRNA组VZ区BrdU+前体细胞的百分比显著高于scr#1 shRNA组(图2a,b)。通过导入DOCK7的cDNA可显著逆转VZ区BrdU+前体细胞的增加(图2a,b)。
该研究进一步通过免疫荧光染色磷酸化的组蛋白H3(PH3)(一种有丝分裂标记),研究VZ区细胞有丝分裂水平(细胞中转染PH3阳性的百分比)。Dock7#2 shRNA组的有丝分裂水平明显高于对照组和逆转组(图2c,d)。因此,敲除DOCK7表达显著促进了VZ区神经前体细胞的增殖。当过表达DOCK7时,可观察到相反的结果(图2e–h)。
为了进一步了解DOCK7表达的改变如何影响神经前体细胞的增殖,在E13.5天进行胚胎电转,2天后对注射BrdU。对进入细胞周期(BrdU + Ki67 +)或退出细胞周期(BrdU + Ki67−)的细胞进行定量(图2i–n)。在Dock7敲除组中BrdU和Ki67双阳性细胞的百分比升高(图2i,j),在DOCK7过表达组中的比例降低(图2l,m)。相反,BrdU+Ki67-细胞的百分比在敲除组降低(图2i,k),在过表达组升高(图2l,n)。这些数据表明DOCK7敲除促使细胞进入细胞周期,而DOCK7过表达则促进退出细胞周期。

图2
3、DOCK7影响神经元形成
根据上述研究结果,DOCK7可能影响RGC从增殖向分化和神经发生的转换。为了证实这一点,在E13.5天胚胎电转DOCK7敲除或过表达的质粒,用Pax6标记RGC,用Tbr2标记VZ基底面的前体细胞。与对照组相比,DOCK7敲除组Pax6+细胞增多而Tbr2+细胞减少(图3a–d)。与之相反,DOCK7过表达可导致Pax6+细胞减少而Tbr2+细胞增多(图3e–h)。因此,抑制DOCK7表达增加RGC细胞池内细胞的数量,而DOCK过表达则减少细胞池数量,并促进RGC分化为基底前体细胞。
E15.5天新生成的神经元通过中间区迁移至皮质板层,而保留自我更新能力的RGC则留在VZ区。与对照组相比,DOCK敲除组的细胞更多分布在VZ区,而在皮质板层和中间区分布较少(图3i,j)。此外还观察到DOCK7敲除组在基底前体细胞分布的SVZ区分布显著减少。此外,Dock7敲除组中,Tuj1阳性细胞的百分比显着降低(图3k)。
当研究DOCK7过表达对神经发生的影响时,本研究观察到VZ区转染细胞的百分比减少,SVZ和中间区域细胞数量增加,但皮质板层的转染细胞数量减少(图3l,m)。DOCK7表达后,Tuj1阳性转染的细胞数量显着增加(图3n);然而,这些细胞大多数位于中间区,而在皮质板层中细胞数量相对较少(图3l,m)。值得注意的是,这些位于中间区的大部分细胞都表现出多极性。DOCK7过表达促进了神经元的形成,该神经元能够到达中间区,但向皮板层迁移时存在缺陷或延迟,这可能是由于神经元从多极性向双极性转变的过程存在缺陷所致。因此,除了控制神经元的发生,DOCK7还可能影响中间区神经元的极化和/或迁移。

图3
4、DOCK7对区间动态核迁移至关重要
影响RGC增殖的因素包括细胞周期时间、细胞极性和不对称分裂,以及最新发现的INM。改变DOCK7的表达水平不会影响细胞周期的时间。然而,在BrdU标记实验中发现,DOCK7敲除组和DOCK7过表达组BrdU标记的细胞核的位置有所不同,提示DOCK7可能影响INM(图2a,e) 。与对照组相比,Dock7敲除组BrdU阳性细胞核的分布更靠近管室面,而在DOCK7过表达组中,细胞核位于VZ区更靠近基底端的位置。为了进一步确定DOCK7是否以及如何影响INM,在E13.5天胚胎电转,2天后注射BrdU,在BrdU注射后15分钟,2小时,4小时和6小时确定转染细胞BrdU标记的细胞核位置(图4)。
在BrdU注射后15分钟,对照组、敲除组及过表达组中,VZ区大多数BrdU标记的细胞核都位于VZ区的上半部分,这与RGC细胞核在S期的已知位置一致(图4a,b)。在对照组中,细胞核在注射后2小时内向室管膜区迁移,并在接下来的4小时继续向室管膜区迁移(图4a,b),与既往研究中S期向G2期转化过程的细胞核迁移方向一致。在细胞核到达室管膜区表面后,RGC进行有丝分裂(距室管膜表面<20um)(图4c,d)。DOCK7敲除加速了细胞核从基底端向室管膜端迁移(图4a)。在注射后2小时,对照组仅一小部分细胞核到达室管膜面,而DOCK敲除组的细胞核的已有很大一部分已经到达该位置(图4a),并在之后的4小时继续存在于该位置,进行有丝分裂(图4c)。实际上,在注射BrdU后6小时而非2小时,在DOCK7敲除组的VZ区中观察到的更多BrdU + PH3 + 细胞图4c)。相反,在检测DOCK7过表达组的VZ区细胞的细胞核位置时,BrdU标记的核从基底端向室管膜面迁移的过程被延迟,在注射后6小时,许多细胞核仍保留在VZ区的上半部分(图4b)。与对照组相比,在DOCK7过表达组中,BrdU + PH3 +细胞在基底端(距室管膜表面> 20 µm)的百分比较高,这表明细胞核向顶端迁移的延迟与细胞远离顶端有丝分裂有关(图4d)。
为了进一步证实上述发现,该研究在胚胎电转后两天进行时差显微成像(图5)。并用胚胎电转技术标记细胞质和细胞膜。切片后进行时差显微成像8至10小时。在对照组中,细胞核稳定地从基底端向室管膜迁移,并在室管膜进行有丝分裂(图5a,e)。而大部分DOCK7过表达细胞的细胞核滞留在基底端,并且细胞远离顶端的位置进行有丝分裂(图5b,e)。这些发现证实了DOCK7敲除会加速细胞核从基底端向顶端迁移的过程,从而导致细胞核在顶端的时间延长,并导致顶端有丝分裂。综合以上结果,可以得出结论,DOCK7严格调节了细胞核从基底端向顶端迁移的过程。

图4


图5
5、DOCK7与TACC3相互作用
以三个不同的DOCK7片段作为引物(图6a)使用双杂交法在胎鼠大脑cDNA库中进行筛查。其中一个片段,DOCK7 506–1164(DOCK7-R2),包含两个阳性克隆,其中一个为与编码Tacc3序列匹配的cDNA,而另一个为包含编码Tacc3全长的cDNA。TACC3是TACC家族的成员,是与中心体和微管相关的蛋白质,与中心体引导的微管形成及核迁移有关,值得注意的是,它还与小鼠皮层发育过程中维持神经前体细胞细胞池数量相关。
使用过表达DOCK7和EGFP-TACC3的人胚胎肾细胞系(HEK293)的裂解物,通过共免疫沉淀实验确认了两者的相互作用(图6b)。GST-TACC3融合蛋白可有效地抑制DOCK7的表达(图6d)。在人SK-N-BE神经母细胞瘤细胞中进行的免疫细胞化学研究表明,DOCK7和TACC3共定位,特别是在中心体位置(图6e)。综上,这两种蛋白存在相互作用。为了进一步研究DOCK7中TACC3结构域的作用,合成了在R2片段缺失的突变体(图6a),并测试了它们与TACC3是否能相互作用。DOCK7中R2片段缺失后,不结合TACC3(图6f)。


图6
6、DOCK7通过拮抗TACC3调控INM和神经发生
敲除TACC可以阻止细胞核从基底端迁移至室管膜面,减少增殖前体细胞的数量并增加神经元的生成,这与DOCK7敲除的作用相反。本研究第一次单独敲除TACC3,结果显示敲除TACC3会抑制细胞核从基底端向顶端迁移,降低BrdU+细胞及有丝分裂细胞百分比,并增加Tuj1 +细胞百分比(图7a-e)。这些数据表明DOCK7和TACC3在皮质神经发生过程中具有相反的功能。
E13.5天胚胎电转敲除DOCK7和/或TACC3的质粒,并在2 d后进行染色。单独敲除DOCK7可导致在室管膜面有丝分裂分裂的细胞百分比更高,VZ区增殖细胞数量增加,神经元生成减少。但是,同时敲除DOCK7和TACC3可完全逆转上述改变(图7f–j)。而为TACC3结合点突变的DOCK7过表达无法逆转DOCK7敲除所致的INM异常,VZ区增殖前体细胞数量增加和神经元生成减少(图7k-o)。总之,这些数据表明DOCK7通过拮抗TACC3功能来调控INM和皮质神经发生。

图7
7、DOCK7拮抗TACC3微管相关功能
TACC蛋白可通过调节与中心体和细胞核偶联的中心体相关微管的生长和完整性来调控皮质神经前体细胞的INM。因此,本研究观察DOCK7是否拮抗TACC3的微管生长促进功能。使用非洲绿猴肾成纤维细胞样细胞系(COS7)进行研究,与TACC3的作用相反,过表达DOCK7减少了微管的体积,值得注意的是,同时过表达DOCK7和TACC3,可使微管的体积恢复至对照载组大小(图8a,b)。为了确定这些操作是否影响微管成核,本研究使用诺考达唑(使微管解聚)处理COS7细胞,并使其在药物洗脱后得以恢复。在洗脱后5分钟,所有组中的大多数转染细胞均显示出清晰的微管结构,表明微管成核不受影响。但是,在洗脱后20分钟,与对照组及共表达DOCK7和TACC3的细胞相比,过表达DOCK7和过TACC3的细胞分别显示出较小和较大的微管,如图8a所示。
在培养的皮质前体细胞中,敲除TACC3显著降低了微管叉状结构的大小,而敲除DOCK7则显著增加了微管叉状结构大小(图8c,d)。值得注意的是,同时敲除TACC3与DOCK7使微管叉状结构的大小恢复到对照组的大小(图8c,d)。
RGC细胞核与中心体的距离受微管生长和完整性改变的影响。胚胎电转后的脑切片用β-微管蛋白(中心体标记)染色,并用DAPI(核标记)复染。TACC3敲除的RGC中,细胞核与中心体的距离增加,与之相反,该距离在DOCK7敲除的RGC中显着减小,并且可以通过同时敲除DOCK7和TACC3来逆转。(图8e,f)。
结合以上数据,这些发现表明,DOCK7拮抗TACC3的微管生长促进或微管稳定功能。值得注意的是,DOCK7的这种作用与其DHR2结构域无关,因为过表达DHR2结合点突变的DOCK7能够逆转敲除DOCK7引起的细胞核-中心体距离减少。
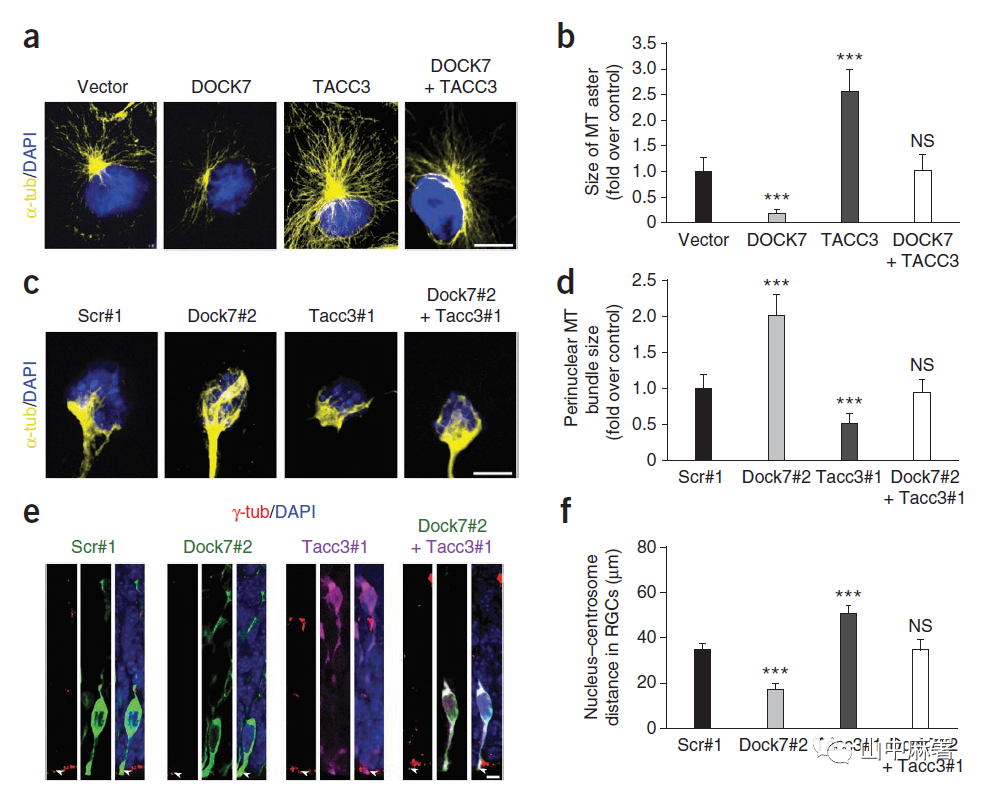
图8
结论
皮层的神经发生依赖于放射状神经胶质细胞(RGC)增殖与分化保持平衡,但如何控制这种平衡的分子机制仍不清楚。本研究提示,DOCK180蛋白质家族的成员,DOCK7,调节RGC的增殖与分化。在小鼠胚胎RGC中,敲除DOCK7可抑制细胞分化,并促进细胞自我更新。与之相反,过表达DOCK7促进RGC分化为基底前体细胞和神经元。本研究进一步提出证据,证明DOCK7通过控制RGC的区间动态核迁移来影响神经发生。DOCK7通过拮抗中心体相关蛋白TACC3的微管生长促进功能发挥作用。因此,DOCK7与TACC3的相互作用,调控了皮层发育过程中的INM和RGC神经发生。
原始文献:
Yang YT, Wang CL, Van Aelst L. DOCK7 interacts with TACC3 to regulate interkinetic nuclear migration and cortical neurogenesis. Nat Neurosci. 2012;15(9):1201-1210. doi:10.1038/nn.3171
作 者: | 刘施雯 |
排 版: | 蒋 明 |
校 审: | 方 芳 |
缪长虹 |


人点赞

人收藏

打赏









 010-82736610
010-82736610
 股票代码: 872612
股票代码: 872612
 京公网安备 11010802020745号
京公网安备 11010802020745号










