


















乳酸在炎症和肿瘤微环境中的免疫调节作用




癌症和免疫介导的炎性疾病(IMIDs),如类风湿性关节炎和多发性硬化症,具有相似的代谢微环境,但两者免疫状态不同。

肿瘤微环境可抑制肿瘤免疫、促肿瘤发生;慢性炎症组织的微环境则促进炎症、延缓炎症消退。尽管这两种环境的免疫学状态大相径庭,然而其代谢状态却相似:两者均为低氧状态,乳酸和其他代谢副产物的水平升高,营养物质含量低。2020年, Certo, M等人在《Nature reviews. Immunology》上发表了一篇题为《Lactate modulation of immune responses in inflammatory versus tumour microenvironments》的综述,描述了与正常组织相比,乳酸在肿瘤和炎症微环境中生物活性的不同,从而有助于疾病中特定免疫状态的建立。现简单介绍如下:

癌症和免疫介导的炎性疾病(IMIDs),如类风湿性关节炎和多发性硬化症,具有相似的代谢微环境,但两者免疫状态不同。肿瘤微环境(TME)中肿瘤细胞和癌症相关成纤维细胞代谢加速共同形成促进肿瘤生长的免疫环境。肿瘤组织耗尽了局部的能量,迫使邻近免疫细胞在缺乏营养的情况下处理乳酸等高浓度的代谢物,导致免疫抑制和肿瘤生长。与TME相比,以关节炎滑膜为例的炎性疾病微环境具有高水平的炎性细胞亚群,例如TH1细胞、TH17细胞、炎性巨噬细胞,以及功能受损的CD4+CD25+Treg细胞,炎症细胞维持IMID的病理过程持续存在。在IMIDs,乳酸由间质成纤维细胞和代谢活跃的浸润性免疫细胞产生,是炎症组织微环境的关键驱动因素。现代观点认为乳酸是细胞代谢的重要碳源和信号分子。TME和IMID微环境中细胞对乳酸的反应截然不同,因此在这一背景下的新研究具有多种临床应用潜力。
乳酸控制免疫反应
下面介绍一些由高乳酸水平调节的代谢途径,其与慢性炎症性疾病和癌症的发生和发展有关。
乳酸脱氢酶
乳酸脱氢酶以两种不同的异构体LDHA和LDHB存在,可以以五种不同的组合方式组装成同四聚体或异四聚体。LDHA将丙酮酸转化为乳酸和NAD+,而LDHB将乳酸转化为丙酮酸,促进氧化代谢。
LDHA通过增加干扰素-γ(IFNG)的乙酰化和转录来促进T细胞效应功能,在炎症中发挥关键作用。细胞毒性T细胞等效应T细胞的增殖和细胞因子的产生高度依赖于糖酵解,因此在低葡萄糖水平和高乳酸浓度条件下变得不活跃。Treg细胞较少依赖糖酵解,而是依赖氧化磷酸化产生能量。因此,乳酸治疗降低效应T细胞功能而不影响Treg细胞功能。抑制LDH后,即使有乳酸存在,也能恢复效应T细胞的功能,这表明乳酸对这些免疫细胞的影响高度依赖LDH。
抑制剂FX11抑制巨噬细胞LDHA活性,可通过抑制丝裂原活化蛋白(MAP)激酶磷酸化下调促炎性细胞因子,诱导型一氧化氮合酶(iNOS)和环氧化酶2(COX2),从而起到抗炎作用。同样的LDH抑制剂可以降低ATP水平,诱导癌症中的氧化应激和细胞死亡。一些研究支持乳酸和LDHA在肿瘤进展中的作用。胰腺癌细胞Pan02在敲除LDHA后,在C57BL/6小鼠成瘤作用受到抑制;在同一模型中,LDHA耗尽的肿瘤组织自然杀伤(NK)细胞显示出更高的杀伤活性,表明LDH活性的增加通过抑制免疫细胞的功能导致肿瘤免疫逃逸。
LDH是一种细胞质酶,但也可以在细胞核中与mRNA结合,这表明LDH在基因表达的转录后修饰中起作用。例如,LDH与编码粒细胞巨噬细胞集落刺激因子(GM-CSF)的RNA富含AU元素结合。因此,LDH可以通过其酶促作用或基因调节功能促进肿瘤的生长。
PKM2
丙酮酸激酶(PK)是一种糖酵解酶,催化磷酸烯醇式丙酮酸不可逆脱磷酸成丙酮酸,产生ATP。在PK的各种同工型中,PKM2因其在免疫细胞和癌细胞中的多种功能而受到特别关注。PKM2的二聚体形式可以将葡萄糖代谢导向生物合成途径,包括磷酸戊糖途径,可以作为激酶进入细胞核调节转录,也可以通过与线粒体外膜结合来维持线粒体功能。
PKM2通过其在Warburg效应中的作用调节免疫细胞的代谢和功能,PKM2在几种炎症疾病中表达增加,如活动性克罗恩病和磺酸诱导结肠炎小鼠的肠道组织中。PKM2的二聚体形式在脂多糖(LPS)激活的巨噬细胞中增加,可以调节HMGB1,进而刺激促炎细胞因子。核PKM2可磷酸化转录因子信号转换器和转录激活因子3 (STAT3),促进IL-6和IL-1β的产生。N,N'-二芳基磺酰胺(DASA)可稳定PKM2四聚体,抑制二聚体PKM2易位至细胞核,减少IL-17的表达。另一种PKM2核易位抑制剂TEPP-46减少TH17细胞极化和实验性自身免疫性脑脊髓炎(EAE)的发生。
在癌细胞中,PKM2二聚体可以易位到细胞核中稳定HIF1α并诱导糖酵解基因的表达。PKM2可与诸多细胞因子相互作用。它与RNA结合蛋白HuR相互作用,调节HuR亚细胞定位、细胞周期进程和胶质瘤细胞生长,并与富含AU蛋白三聚氰胺(TTP)相互作用。通过蛋白酶体降解破坏TTP的稳定性,以调节乳腺癌细胞的增殖。PKM2可进入到线粒体中,使BCL-2磷酸化和稳定化,促进癌细胞适应氧化应激。PKM2与细胞核中的NF-κB和HIF1α的相互作用是肿瘤生长过程中VEGF-A分泌增加和血管形成的原因。PKM2通过诱导趋化因子,如CCL8、CCL2和CXCL1的释放,促进肿瘤相关巨噬细胞和髓源性抑制细胞向TME的募集。这些细胞通过诱导Treg细胞和抑制NK细胞的功能来发挥免疫抑制功能。二聚体PKM2还可以通过与PDL1启动子中的缺氧反应元件结合,调节肿瘤相关免疫细胞中PDL1的表达,从而创造免疫逃避肿瘤环境。
MAVS
线粒体抗病毒信号蛋白(MAVS)介导NF-κB和干扰素调节因子3(IRF3)的活化以应对病毒感染,随后I型干扰素如IFNβ表达。导致干扰素产生的级联反应始于先天免疫受体的激活,包括视黄酸诱导基因I (RIG- I)样受体和Toll样受体。MAVS与乳酸直接结合而失活,抑制了RIG-I样受体的信号激活和I型干扰素的产生。MAVS通过己糖激酶2(HK2)与线粒体结合,促进糖酵解途径。RIG-1样受体激活后,MAVS与HK2的结合减少,与RIG-1的结合增加,使HK2从线粒体释放并失活。乳酸使MAVS不再能够与线粒体结合,RIG-I-MAVS复合物的形成和干扰素的产生受到抑制。鉴于I型干扰素在调节免疫细胞(如树突状细胞,Treg细胞和细胞毒性T细胞)中发挥着关键作用,这可能是乳酸促进TME中免疫监视抑制的一种机制。
在慢性炎症的情况下,乳酸处理使CD4+T细胞线粒体表面HK2结合位点增加。HK2与线粒体外膜的阴离子通道电压依赖性结合可促进细胞存活,这可以解释浸润慢性炎症组织(如类风湿关节炎滑膜)的T细胞的持续存在和存活。
脂肪酸合成
脂肪酸合成(FAS)在调节免疫细胞的活性中起关键作用。免疫细胞活化后能量主要来自于糖酵解,导致丙酮酸水平升高,产生并分泌大量乳酸。丙酮酸也可以在线粒体内转化为乙酰辅酶A和柠檬酸,柠檬酸盐输出到细胞质中,是FAS的主要原料。
脂质代谢调节巨噬细胞向炎症表型极化,脂肪酸氧化或糖异生促进了M2巨噬细胞的抗炎功能。但在M1巨噬细胞中,糖酵解的增加是至关重要的,不仅可以更快地产生ATP,还可以通过三羧酸(TCA)循环获得柠檬酸,以维持新生FAS。在饮食诱导的小鼠炎症模型中,敲除脂肪酸合酶(Fasn)导致募集到脂肪组织中的巨噬细胞减少。
在T细胞活化过程中,调控脂肪酸生物合成的基因上调。乙酰辅酶A羧化酶1(ACC1)和FASN活性上调增加了TH17细胞的致病性。ACC1的抑制剂Soraphen A可阻断CD4+T细胞分化为TH17细胞,有利于T细胞分化为FOXP3+Treg细胞。该机制可降低EAE的严重程度。类似浓度的乳酸使CD4+T细胞上调脂肪酸的从头合成,导致IL-17的生成增加和细胞活力降低。
正常细胞主要依赖脂肪酸的摄取和氧化,然而在肿瘤细胞中,即使在微环境中存在脂质,也会重新激活FAS。这表明该途径在它们的代谢中起着关键作用。癌细胞中激活的FAS会改变其膜成分和流动性,变得更耐化疗。免疫细胞中FAS的激活也会导致TME中这些细胞功能的改变,例如肿瘤树突状细胞的免疫刺激能力降低。鉴于FAS在炎症疾病和癌症中调节多个细胞过程,这一代谢途径的调节为治疗带来了巨大的希望。
乳酸穿梭功能
乳酸在产能和耗能细胞之间穿梭,在生理上起着至关重要的作用,包括作为重要
的能量来源、糖异生前体和信号分子。例如,在骨骼肌中,快肌纤维糖酵解并产生乳酸,然后进入慢肌纤维细胞。同样,在大脑神经元胶质细胞代谢耦合中,星形胶质细胞通过糖酵解输出乳酸作为相邻神经元的能量来源。乳酸从糖酵解细胞(快收缩纤维和星形胶质细胞)到氧化细胞(慢收缩纤维和神经元)的载体运输部分是由溶质载体的专门运输(称为乳酸穿梭)提供的。乳酸在炎症性疾病微环境和TMEs中的各种功能,通常是对立的。

图1 乳酸在炎症性疾病微环境中的免疫调节—以关节炎滑膜为例
乳酸在炎症性疾病微环境的免疫调节
在炎性疾病的背景下,乳酸触发一系列胞内信号,促进慢性炎性过程。乳酸介导的对延迟低氧的反应似乎与HIF1α不偶合,因为它使用NDRG家族成员3(NDRG3)。NDRG3在常氧状态下以PHD2/VHL依赖的方式降解,类似于HIF1α。但是,在长期缺氧条件下,它通过与乳酸结合而免于降解。NDRG3水平的升高导致RAF-ERK信号通路的激活,调控缺氧相关的病理生理反应,包括炎症和血管生成。LDHA的药理或遗传抑制以剂量依赖的方式限制NDRG3蛋白的积累。
在关节炎滑膜中,乳酸通过多种方式调节免疫细胞的功能,即迁移和细胞因子的产生(图1)。在T细胞中,乳酸通过CD4+T细胞乳酸转运蛋白SLC5A12和CD8+T细胞的SLC16A1(MCT1)诱导“停止迁移”信号,使它们在炎症部位积累。炎症组织中乳酸介导的T细胞运动抑制与糖酵解减少相结合。乳酸钠抑制CD4+T细胞的几种糖酵解酶和葡萄糖流量表达减少,使T细胞在炎症部位积累。类风湿关节炎中初始CD4+T细胞由于6-磷酸果糖-2-激酶/果糖-2,6-二磷酸酶(PFKFB3)下调抑制了细胞内糖酵解水平,导致6-磷酸葡萄糖(G6P)向磷酸戊糖途径分流,产生NADPH,改变了对毛细血管扩张性共济失调症突变蛋白(ATM)的激活,ATM是一种参与细胞周期调节的重要酶。总之,这些变化导致类风湿性关节炎中CD4+T细胞增殖活跃并产生TH1、TH17等促炎亚细胞群,导致慢性炎症。
类风湿性关节炎中的ATPlowpyruvatelowNADPHhiCD4+T细胞也表现出FAS增强,导致细胞质脂滴积聚和TKS5上调。因此类风湿性关节炎患者的TKS5hiCD4+T细胞形成富含肌动蛋白和皮质蛋白的膜皱褶,增加了浸润炎症组织的能力。抑制FAS可以逆转以上所有作用。FAS不仅导致了炎症部位CD4+T细胞浸润增加,而且还导致了它们在该部位的滞留。
在关节炎滑膜中,乳酸或SLC5A12诱导的特定代谢途径的激活促进CD4+T细胞浸润并分化为TH17细胞和T滤泡辅助细胞亚群,在同一炎症环境中诱导形成不同的淋巴结构,在类风湿性关节炎的发病机理中起着至关重要的作用。
在小鼠关节炎模型中,抑制SLC5A12减少了乳酸的摄取,使T细胞功能恢复并减轻了炎症反应,这表明它可能是治疗IMIDs炎症的新靶点。滑膜细胞代谢需求的升高导致类风湿性关节炎患者滑液中乳酸的积累,滑液乳酸水平是区分炎症性关节炎和脓毒性关节炎的可靠指标。与骨关节炎患者相比,类风湿关节炎患者的血清和滑液中LDH同工酶含量更高,滑膜组织中的LDH活性较健康对照组增强。乳酸可以激活Gi蛋白偶联受体81 (GPR81),抑制cAMP和蛋白激酶A (PKA)信号转导和β-抑制蛋白的活化。通过在单核细胞和巨噬细胞中激活这一机制,乳酸可以减少胰腺炎和肝炎模型中的炎症。
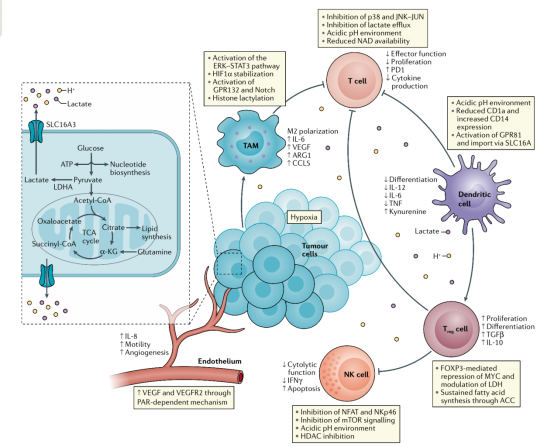
图2:乳酸在TME中的免疫调节
乳酸在TME中的免疫调节
1923年,Otto Warburg首次观察到癌细胞的有氧糖酵解,即使在有足够氧气的情况下,也表现出高糖摄取和过量乳酸形成,被称为Warburg效应,至今仍是癌症的十大特征之一。癌细胞产生的乳酸分泌到细胞外环境中,促进癌症进展。癌细胞或基质细胞中与质子偶联的乳酸外流可调节TME(包括细胞侵袭、血管生成、生存信号、转移发展和逃避免疫监视)来促进肿瘤进展。细胞外酸中毒抑制T细胞介导的免疫,中和肿瘤酸度可提高对免疫疗法的抗肿瘤效应。细胞外pH值在6.0–6.5之间诱导肿瘤特异性CD8+T细胞呈无能状态,其细胞溶解活性和分泌细胞因子的能力降低,而质子泵抑制剂治疗可有效恢复T细胞功能。TME中低pH降低了M1巨噬细胞中iNOS、CCL2和IL-6的表达,增加M2巨噬细胞标志物的表达。细胞外酸化也通过mTOR抑制NK细胞的抗肿瘤活性。
TME中癌细胞、T细胞、NK细胞、树突状细胞和巨噬细胞等细胞可以感知胞外乳酸水平,从而触发细胞内信号,微调细胞行为并强烈影响它们的功能(图2)。乳酸诱导的GPR81活化参与肿瘤生长,调节乳酸摄取和代谢相关基因的表达。乳酸可以激活巨噬细胞中的G蛋白偶联受体132(GPR132),促进巨噬细胞向M2极化。来自乳酸的乳酰基可用于组蛋白的翻译后修饰,提高IL6和ARG1基因的表达。乳酸抑制单核细胞分化成树突状细胞,抑制T细胞产生细胞因子和细胞毒性活性。TME中的乳酸降低NK细胞的溶细胞功能并增加髓样抑制细胞的数量。乳酸可阻止T细胞和NK细胞中NFAT的活化,使IFNγ产生减少,这与乳酸在炎性环境中促进IFNγ产生的作用相反。
LDHA的表达受HIF1α、MYC和p53调节,可促进上皮-间质转化,血管生成和侵袭增加。LDHB表达下调或缺失是前列腺癌、乳腺癌和胰腺癌等癌症发展中一个重要的早期事件,与高增殖、易浸润和患者的不良预后相关。在葡萄糖缺乏的情况下,LDHB介导的乳酸使用支持自噬以维持新陈代谢和癌细胞的生长。LDHB表达的变化往往与早期代谢适应有关,表明乳酸的产生和使用可能参与了肿瘤细胞的代谢适应,以支持转移。骨转移性乳腺癌细胞释放大量乳酸。乳酸是破骨细胞的关键能量来源,嗜骨性肿瘤细胞可释放乳酸促进破骨细胞的分化和代谢重编程,允许肿瘤细胞侵袭转移灶。乳酸刺激内皮细胞释放VEGF促进伤口愈合和肿瘤相关的血管生成。在胶质瘤细胞中,乳酸诱导转化生长因子β2的表达,转化生长因子β2是癌细胞迁移、侵袭、上皮-间质转化和转移灶形成的关键调控因子。综上所述,乳酸和LDH的表达可以支持肿瘤细胞的代谢适应和肿瘤发生。
“论肿道麻”点评
癌症和IMIDs具有相似的代谢微环境但两者免疫状态不同。TME中肿瘤细胞和癌症相关成纤维细胞代谢加速共同形成促进肿瘤生长的免疫环境。肿瘤组织耗尽了局部的能量,迫使邻近免疫细胞在缺乏营养的情况下处理乳酸等高浓度的代谢物,导致免疫抑制和肿瘤生长。与TME相比,以关节炎滑膜为例的炎性疾病微环境具有高水平的炎性细胞亚群,例如TH1细胞、TH17细胞、炎性巨噬细胞,以及功能受损的CD4+CD25+Treg细胞,炎症细胞维持IMID的病理过程持续存在。在IMIDs,乳酸由间质成纤维细胞和代谢活跃的浸润性免疫细胞产生,是炎症组织微环境的关键驱动因素。现代观点认为乳酸是细胞代谢的重要碳源和信号分子。TME和IMID微环境中细胞对乳酸的反应截然不同,因此在这一背景下的新研究具有多种临床应用潜力。
了解在癌症和IMIDs的患病组织中局部代谢物控制免疫应答的机制具有巨大的治疗意义。深入的基础研究是为了更好的临床医疗。靶向特定的代谢途径正在成为IMIDs中有用且有前途的治疗策略,并且未来在免疫代谢方面的研究有望提供可以靶向调节免疫细胞活性且副作用更少的新药。
审校:张军,缪长虹
原始文献:Lactate modulation of immune responses in inflammatory versus tumour microenvironments. Certo M, Tsai CH, Pucino V. et al. Nat Rev Immunol, 2021; 21(3):151-161. doi: 10.1038/s41577-020-0406-2

不感兴趣
看过了
取消

人点赞

人收藏

打赏
不感兴趣
看过了
取消













 010-82736610
010-82736610
 股票代码: 872612
股票代码: 872612
 京公网安备 11010802020745号
京公网安备 11010802020745号










