
本研究利用阿拉伯糖(ara)诱导的pBAD启动子调控大肠杆菌的合成代谢途径。
编译:微科盟艾奥里亚,编辑:微科盟汤貝、江舜尧。
微科盟原创微文,欢迎转发转载,转载须注明来源《微生态》公众号。
论文ID
原名:Metabolome and proteome analyses reveal transcriptional misregulation in glycolysis of engineered E. coli
译名:代谢组和蛋白质组分析揭示了工程大肠杆菌糖酵解过程中的转录失调
期刊:Nature Communications
IF:14.919
发表时间:2021.8.13
通讯作者:Hannes Link
通讯作者单位:德国马克斯·普朗克陆地微生物研究所
实验设计
结果
为了探究诱导合成代谢途径如何对宿主代谢造成影响,我们在大肠杆菌中表达了酵母的甘油生物合成途径(图1a)。甘油途径共涉及两步途径,起始于糖酵解的代谢产物磷酸二羟丙酮(DHAP)。3-磷酸甘油脱氢酶1(GPD1)催化DHAP转化为3-磷酸甘油(甘油-P),随后3磷酸甘油水解酶2(GPP2)催化甘油-P去磷酸化并形成终产物甘油。本研究中,我们将能够表达编码gpd1和gpp2两个基因的、同时缺乏甘油激酶基因(glpK)来防止甘油被重新作为碳源而利用的质粒导入到大肠杆菌中,构建了能够产甘油的大肠杆菌菌株,并将其定义为基础菌株。
我们试图通过表达甘油途径中具有阿拉伯糖诱导的pBAD启动子的第一种酶(GDP1)和具有强构型促进作用的第二种酶(GPP2)来调控甘油的产生(图1a)。用pBAD启动子表达的GFP表现出在阿拉伯糖(ara)浓度和启动子活性之间存在一种线性关系(图1b)。因此,我们预期pBAD启动子将允许我们线性控制GPD1的丰度,从而逐渐增加甘油的产生(图1c)。然而,较低浓度的ara(0.3%)导致强烈的生长缺陷和较低的生物量浓度和甘油滴度(图1d)。0.1%的ara浓度(17.71 mM,图1d)可达到最大甘油滴度。
随后,我们对较高ara浓度下引起的生长负担的机制进行了探究。由于pBAD启动子表达的GFP并不影响细菌的生长,因此我们排除了GPD1蛋白表达所造成的影响。因此,生长负担可能是由甘油和糖酵解通量之间的竞争引起的。基于大肠杆菌代谢的基因组尺度上的模型进行通量平衡分析(FBA),我们预测细菌生长和甘油生成速率遵循线性关系(图1e中的直线),这反映了利用葡萄糖生产生物量或甘油之间的权衡。为了测试基础菌株是否遵循这一理论权衡,我们测量了三个诱导水平下(0、0.1和0.5%)的甘油生成率和生长速率(图1e中的点)。然而,实验测定的速率不符合FBA预测的理论权衡(图1e)。在0.5% ara时测得的甘油生成率和生长率明显低于理论生成率和生长率,因此表明还存在除通量平衡以外的其他因素。
总之,pBAD启动子使我们能够线性增加蛋白表达(图1b)。然而,根据通量平衡分析估计的理论权衡,我们不能使用pBAD启动子来调节生长速率和甘油生成速率(图1e中的线)。相反,在0.5%的甘油浓度诱导下,测量的生长速率下降比FBA预测的更强(图1e)。

为了了解引起基础菌株生长负荷的分子机制,我们在三个ara浓度诱导下(0、0.1和0.5%),对大肠杆菌的代谢组进行了测定。细菌菌株在摇瓶中培养后,通过快速过滤的方法收集用于代谢组学的样品(图2a)。代谢组数据涵盖96种代谢物,其在0.1% ara浓度下保持相对恒定,但在0.5% ara浓度时显示出强烈变化(图2b)。在0.5% ara浓度时下降最显著的代谢物是甘油途径的直接前体DHAP(图2b)。此外,直接位于DHAP上游的1,6-二磷酸果糖(FBP)是反应最强烈的代谢物之一,其在0.5% ara浓度下下降5倍以上(图2b)。这些数据表明,甘油过量生产扰乱了工程菌代谢起始步骤附近的代谢物。
FBP作为一种调节代谢物,其负责大肠杆菌中糖酵解通量依赖性基因表达的调节。FBP抑制转录因子Cra的活性,该转录因子能够抑制糖酵解酶编码基因的表达并激活糖异生相关基因(图2c)。尽管目前尚不清楚FBP是Cra的直接效应因子还是间接效应因子,但人们普遍认为FBP的浓度会影响Cra的活性。相应地,我们进一步探究了低浓度的FBP(在0.5% ara浓度背景条件下)是否会激活Cra转录因子,从而改变基因表达和酶水平。我们在三个诱导水平下对蛋白质组进行测定,并探究了糖酵解和糖异生中共计38种酶的丰度(图2d)。与代谢产物相似,0.5%诱导时的酶水平变化明显强于0.1%诱导时的酶水平变化。为了检测蛋白质组变化是否由Cra引起,我们测量了Cra缺陷株(Δcra)的蛋白质组作为参考(图2d)。磷酸烯醇式丙酮酸合成酶(PpsA)是Δcra菌株中下降最强烈的酶之一。Cra对PpsA表达的强烈影响与之前的研究一致,这表明PpsA启动子受Cra的直接控制。在我们的基础菌株中,PpsA是在0.5% ara浓度下增加最强烈的酶之一(图2d),这表明该菌株中的高Cra活性。此外,基础菌株具有被Cra抑制的低水平的糖酵解酶,如3磷酸-甘油醛脱氢酶(GapA)。
综上所述,蛋白质组和代谢组数据表明,0.5%的ara诱导的甘油代谢途径可降低FBP的浓度。这反过来激活了转录因子Cra,然后下调糖酵解中的酶(例如GapA),上调糖异生中的酶(例如PpsA)。由于细胞在最低含量的葡萄糖培养基中生长,因此我们假设糖异生的激活是生长负荷的原因。我们通过删除基础菌株中的cra基因来验证这一假设(图2e)。在合成甘油途径的高诱导下,得到的Δcra菌株确实比基础菌株生长更好,最大甘油滴度增加了1.6倍(比较图2e和1d)。因此,Cra-调控有助于大肠杆菌甘油过量产生的生长负担。

为了获得额外的证据证明Cra的转录调控是甘油生成所产生的问题,我们开发了一个小的动力学模型(图3a)。该模型包括一种代谢物(FBP)和两种酶--e1和e2。酶e1对应于低水平糖酵解中的3-磷酸甘油醛脱氢酶(GapA),e2代表甘油途径中的GPD1。基于Michaelis-Menten动力学(酶反应的公认动力学形式),FBP受低水平的糖酵解(rlower_glycolysis)和甘油途径(rglycerol)反应速率的影响。与通量平衡分析相似(图1e),我们假设模型中的内流恒定,并将糖酵解上层的反应速率固定为4.9 mmol g-1 h-1。这意味着FBP以恒定的速率产生,并根据方程中的质量平衡,其要么用于甘油的生成要么用于生物的生长。
总之,我们分析了三种具有不同调节结构的模型(图3b)。基础菌株的模型包括相互作用,其中FBP激活酶e1的表达。这种相互作用类似于Cra对较低糖酵解的转录调控。有关于Δcra菌株的第二个模型没有调节作用。在第三个模型中,FBP激活酶e1和e2的表达。在这种双重调节模型(2xcra模型)中,较低的糖酵解和甘油途径均受到Cra-调控。我们用影响最大酶表达率的幂律项模拟Cra-调控。由于幂律项在非诱导状态下等于1,因此所有模型具有相同的参数集。
基于5000个参数集对三个模型进行分析,从具有生理学意义的范围内随机进行抽样(表1)。我们对幂律指数在1和2之间进行抽样,以确保Cra-调控至少是线性地依赖于FBP的浓度,并避免指数 < 2时可能发生的不稳定性。对于5000个参数集中的每一个,我们计算了最大甘油生成速率(rglycerol,MAX)--通过给定特定的参数得以实现。为了估计rglycerol,MAX,我们使用了数值延拓法,该方法迭代地增加了酶e2(β2)的表达率,并计算FBP,e1和e2新的稳态。每次迭代后,延拓方法通过检查雅可比矩阵(Jacobian matrix)的特征值来确定模型的稳定性,如果模型中发生不稳定,则终止。如果模型保持稳定,继续方法直到e2达到最大表达率(β2,max)。因此,rglycerol,MAX是延拓法终止点时的甘油生成率,我们获得了三种模型各自的5000个rglycerol,MAX。
5000个 rglycerol,MAX值的分布显示,考虑到Δcra模型中达到最大甘油生成速率(rglycerol,MAX)的参数要多于其在基础菌株模型中的参数(图3c),因此Δcra模型的表现优于基础菌株模型。这与Δcra菌株表现优于基础菌株的实验观察结果相匹配。基本假设是:参数集达到高甘油通量越多,真实系统中达到相同程度的可能性就越高。由于基础菌株模型在较高诱导水平下不稳定(图3c),因此其未达到较高的甘油生成率。为了更好地理解这些不稳定的起源,我们用三个模型和一个平均参数集进行了时程模拟。时程模拟与数值延拓法得到的结果相匹配,从而证实两种数值方法得到的结果相同。我们模拟了不同诱导水平的模型,由于酶e2在高诱导下呈指数增加,因此基础模型在较高诱导下并不稳定。因此,存在一个临界点,即e2的表达率超过其生长稀释度时。由于Cra下调了e1,从而用于生长,因此这些不平衡可能被Cra的调节放大。相比之下,Δcra模型在几乎所有诱导水平均保持稳定。我们分析中的最佳模型是2xcra模型。使用该模型,参数集的最高部分达到了较高的甘油通量(图3c)。此外,2xcra模型的稳定性与Δcra模型相似(图3c)。

为了改造被Cra抑制的pBAD启动子,我们在启动子区域和核糖体结合位点之间插入了与Cra一致的结合序列(图3d)。然后我们在该pBAD-Cra启动子的调控下表达GPD1,并将质粒导入大肠杆菌ΔglpK中,构建了pBAD-Cra菌株。该菌株的长势确实远优于基础菌株,并且即使在2%阿拉伯糖完全诱导下仍保持良好的生长(图3e中的橙色菌株)。在0.5% ara浓度下,pBAD-Cra菌株的生长和甘油生成速率甚至高于理论模型中的临界速率(图3f)。这些数据证实了模型预测,即双Cra-调节菌株的性能优于基础菌株和Δcra菌株。
进一步,我们在本研究所涉及的生产系统背景下比较了pBAD启动子和pBAD-Cra启动子的活性。在0.5% ara浓度条件下,pBAD-Cra启动子的活性比pBAD启动子低3.7倍,在一结果表明Cra-结合位点的插入降低了GPD1的表达。流式细胞仪数据显示,GPD1-GFP含量的细胞间变异与启动子无关,这表明pBAD-Cra群体的所有细胞具有较低的启动子活性。由于Cra能够抑制启动子活性,这导致了pBAD-Cra启动子的活性低于pBAD启动子。理论上,通过其他降低pBAD启动子活性的突变可以实现相同的效果(如-10和-35 boxes之间的突变)。因此,我们构建了一个在-10和-35 boxes之间的pBAD启动子突变体,使活性下降2倍,并对所构建的弱pBAD菌株进行分析。弱pBAD菌株的生长确实优于基础菌株,并能够获得更高的甘油滴度(图4a)。但由于pBAD-Cra和pBAD-weak启动子的活性不同(活性分别比原始pBAD启动子低3.7倍和2倍),这使得弱pBAD菌株并没有优于pBAD-Cra菌株。除此之外,pBAD-Cra菌株存在动态的pBAD-Cra启动子,而弱pBAD菌株的启动子是静息的。
如果Cra主动抑制pBAD-Cra启动子,我们预期在启动子区域外插入Cra-结合位点不会对其造成影响。事实上,只有当结合位点直接插入pBAD启动子后,插入1-3个结合位点才能改善菌株的生长(图4b)。为了进一步证明pBAD-Cra启动子并没有完全失活,我们在野生型和Δcra菌株中采用GFP测定了pBAD启动子和pBAD-Cra启动子的活性。在野生型中,pBAD-Cra启动子的活性低于pBAD启动子(图4c),因此表明Cra抑制了启动子活性。然而,在Δcra菌株中,pBAD-cra启动子的活性略高于pBAD启动子(图4c)。这些结果表明pBAD-Cra启动子是有功能性的:Cra能够抑制pBAD-Cra的启动子,但这种调节作用在缺失Cra的情况下(在Δcra菌株中)无法进行。因此,pBAD-Cra所表现出的低活性不仅是由于序列的变化,同时也是由于Cra的主动抑制效果。
为了进一步证明这种方法的普适性,我们将Cra插入到组成型启动子和pTet启动子中。在这两种情况下,具有Cra调节性的启动子突变体菌株比具有野生菌组生长的更好。这表明Cra抑制这些启动子并自动降低其活性。

为了更好地理解Cra-调节的动态性质,我们测量了甘油途径诱导后代谢组和蛋白质组的动态变化。因此,我们在0.5% ara浓度条件下,诱导了基础菌株和pBAD-Cra菌株,并收集了诱导4.5 h的代谢组学和蛋白质组学样本。此外,我们对培养过程中的菌株生长过程(图5a)和培养基中甘油浓度进行了测定,以计算通过甘油途径的流量(图5b)。通过记录菌株的生长过程,我们同样发现pBAD-Cra菌株的长势要好于基础菌株(图5a)。在诱导1 h后,基础菌株出现了生长缺陷,但在该时间点,两种菌株具有相似的甘油生成率(~5 mmol g-1 h-1,图5b)。诱导2 h后,pBAD-Cra菌株的甘油生成率(10 mmol g-1 h-1)要高于基础菌株(6 mmol g-1 h-1)。这表明菌株生长受到抑制并不是由于甘油的生成所导致,而是由于诸如GPD1的高表达在内的其他效应所引起的(图5c)。
随后,我们假设在诱导1h后,Cra在基础菌株中被激活,而Cra在pBAD-Cra菌株中的活性仍较低。为了验证这一假设,我们再次使用PpsA的丰度作为Cra活性的代表。在诱导1h后,基础菌株中PpsA水平增加,这与该菌株显示生长缺陷的时间相匹配(图5c)。然而,在pBAD-Cra菌株中,其PpsA水平保持恒定,表明在该菌株中,Cra的活性保持在激活糖异生的阈值以下。
如果Cra在pBAD-Cra菌株中的活性低于其在基础菌株中的活性,我们猜测基础菌株中的FBP浓度降低。事实上,基础菌株经过诱导后,其FBP浓度迅速下降(图5d)。相反,尽管pBAD-Cra菌株中经甘油途径中的通量较高,但其FBP浓度始终高于基础菌株。pBAD-Cra菌株中其他糖酵解代谢物(DHAP和PEP等)的浓度也高于基础菌株(图5d)。此外,在0、0.1%和0.5%的ara恒定诱导pBAD-Cra菌株的条件下,基于代谢组学我们证实了pBAD-Cra菌株在稳态条件下维持较高浓度的FBP。
因此,与基础菌株相比,pBAD-Cra菌株可以在更高的甘油生成速率下维持更高的FBP水平。这表明FBP和Cra之间存在相互作用,Cra与pBAD启动子结合,抵消了FBP的降低:(i)如果FBP低于临界值,则Cra的活性增加,(ii)在较高的Cra活性条件下,又会抑制pBAD-Cra启动子的活性,从而降低GPD1的表达,(iii)GDP1的低表达将进一步恢复FBP的浓度。因此,我们的数据表明这种反馈调节使高FBP水平与高甘油生成率的并存成为可能,这可能阻止了大肠杆菌将代谢过程从糖酵解转变为糖异生。然而,这种反馈回路如何在空间和时间上对代谢造成影响还需要进一步的实验来证明。

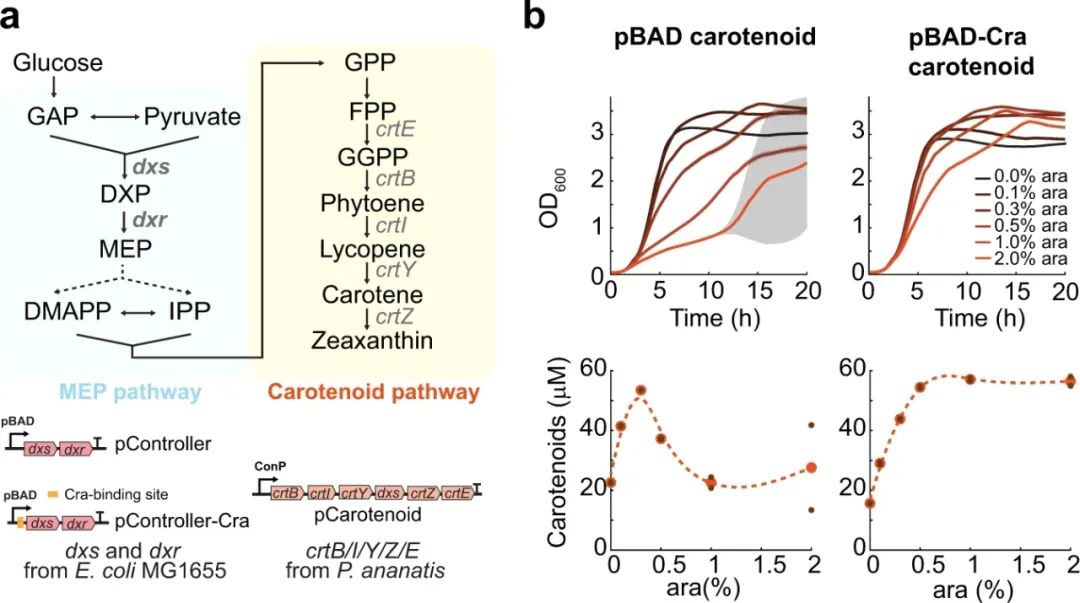
由于许多基于生物的化学物质大多是使用糖酵解的代谢产物作为其合成前体,我们想知道Cra调节的pBAD启动子是否可以具有更广泛的适用性。因此,我们利用pBAD启动子来调控类胡萝卜素过量生产的合成代谢途径(图6a)。类胡萝卜素的生物合成始于糖酵解代谢产物丙酮酸和3-磷酸甘油醛(GAP),随后经MEP途径转化为法尼基二磷酸(FPP),FPP在内生菌(Pantoea ananatis)异源酶的催化下进一步转化生成类胡萝卜素(图6a)。MEP通路中的前两种酶(Dxs和Dxr)使用两种不同的pBAD启动子在质粒中过表达,我们将两种质粒分别称为pController和pController-Cra。类胡萝卜素途径中的其余酶(CrtE/B/I/Y/Z)使用P. ananatis的天然启动子(pCarotenoid)在第二个质粒中表达。
仅pCarotenoid质粒表达时会导致类胡萝卜素的生成,但这并不影响细胞的生长。当大肠杆菌携带pCarotenoid质粒同时与pController或pController-Cra质粒共存时,类胡萝卜素产量增加了近3倍(图6b)。然而,对于与pController质粒共存而言,其仅能在低浓度ara下达到较高的类胡萝卜素浓度,而对于与pController-Cra质粒共存情况而言,其能耐受更广泛的ara浓度。因此,与甘油途径相似,我们观察到具有Cra调控的启动子能够在高诱导水平下维持较高的生长速率,并且高诱导浓度不会影响细胞的生长和生产力(图6b)。综上所述,本研究结果表明Cra-调控的pBAD启动子具有普适性规律,能够赋予生物工程师这样一种能力:调控利用糖酵解代谢物的一系列合成途径的表达。
讨论
本研究利用阿拉伯糖(ara)诱导的pBAD启动子调控大肠杆菌的合成代谢途径。而在GFP表达的情况下,pBAD启动子在诱导剂(ara)浓度和表达率之间表现出线性关系,但这种现象在甘油生产过量时并没有观察到。在甘油生产过量的条件下,诱导剂的小幅增加足以引起生长负担并导致生产力受限。这一现象的结果就是细菌生物量的浓度较低,以及培养结束时甘油滴度较低。
传统的观点认为,合成代谢途径竞争性地消耗了细菌生长资源,因此被视为是工业微生物的负担。最近的多组学数据表明,合成代谢途径可以通过扰乱蛋白质组和代谢组,进而为细胞代谢带来额外的负担。代谢组能够通过改变调节代谢物的浓度进一步调控代谢物的表达和功能,因此蛋白质组的紊乱对于生物体而言至关重要。本研究中,我们观察到甘油的过量生产降低了糖酵解代谢物的浓度,并引起了转录水平上的错误调节。具体而言,低浓度的调节代谢物FBP激活转录因子Cra,从而上调糖异生酶(如PpsA)的表达。这些结果证明了在工程微生物中维持调节代谢物高于临界阈值的重要性。然而,在固有的合成途径中往往缺乏维持代谢物浓度稳态的调节机制,如酶的变构调节或转录调节。本研究中,我们证明了在合成甘油途径中的工程转录调节(Cra-regulation)可以帮助将调节代谢物维持在临界阈值以上:pBAD-Cra菌株比基础菌株具有更高的FBP水平,同时具有更高的甘油通量。这支持了这样一种假说:Cra的依赖性调节通过下调甘油通路的表达来应对FBP水平的下降,从而抵消FBP浓度的下降。然而,由于这种调控是否真实的动态存在仍然是一个悬而未决的问题,我们仍不能排除这样的可能性:由于Cra的持续抑制活性,pBAD-Cra启动子只是作为一个较弱的pBAD启动子发挥作用。有关于pBAD-Cra启动子是否能够自动适应新的环境条件,在未来的研究中应该进一步得到阐述。
之前的研究主要集中在启动子工程的转录结果,或证明工程启动子能够增加合成通路的生产力。本研究中,我们结合代谢组学和蛋白质组学,在宿主代谢水平上探究了工程启动子所产生的结果。从生物技术的角度来看,这种方法将有助于对生产菌株进行改进,使其能够自主缓冲工业规模生物反应器的外部扰动,以及诸如基因表达噪声这样的内部扰动。
不感兴趣
看过了
取消

人点赞

人收藏

打赏
不感兴趣
看过了
取消
©2012-2023 北京华媒康讯信息技术股份有限公司 All Rights Reserved. 注册地址:北京 联系电话:010-82736610
广播电视节目制作经营许可证 —(京)字第 17437号 京海食药监械经营备20200522号
京ICP备12011723号 京ICP证150092号
 京公网安备 11010802020745号
工商备案公示信息
互联网药品信息服务资格证书((京)-非经营性-2020-0015)
京公网安备 11010802020745号
工商备案公示信息
互联网药品信息服务资格证书((京)-非经营性-2020-0015)






您已认证成功,可享专属会员优惠,买1年送3个月!
开通会员,资料、课程、直播、报告等海量内容免费看!

打赏金额
认可我就打赏我~
1元 5元 10元 20元 50元 其它
打赏作者
认可我就打赏我~
扫描二维码
立即打赏给Ta吧!
温馨提示:仅支持微信支付!
已收到您的咨询诉求 我们会尽快联系您


































 010-82736610
010-82736610
 股票代码: 872612
股票代码: 872612




