
HFD通过诱导小鼠肠道微生物失调、代谢改变和肠道上皮屏障功能受损来推动结直肠癌的发生
编译:微科盟牛魔王,编辑:微科盟茗溪、江舜尧。
微科盟原创微文,欢迎转发转载,转载须注明来源《微生态》公众号。
背景与目的:膳食脂肪摄入和结直肠癌风险增加息息相关。我们研究了高脂肪饮食 (HFD) 通过调节肠道微生物群和代谢物驱动结直肠癌的发生。
方法:在偶氮甲烷(AOM)和Apcmin/+结直肠癌模型的小鼠中,将高脂肪饮食(HFD)或对照饮食 (CD) 饲喂给同窝小鼠,同时使用或者不使用抗生素鸡尾酒治疗。然后使用粪便菌群移植的无菌小鼠进行验证并通过宏基因组测序和高效液相色谱-质谱分别检测肠道微生物群和代谢物。最后通过脂多糖 (LPS) 水平和透射电镜测定肠道屏障功能。
结论:HFD通过诱导小鼠肠道微生物失调、LPA升高的代谢紊乱和肠道屏障功能障碍来促进结直肠癌的发生。
论文ID
原名:High-Fat Diet Promotes Colorectal Tumorigenesis through Modulating Gut Microbiota and Metabolites
译名:高脂肪饮食通过调节肠道微生物群和代谢物促进结直肠癌发生
期刊:Gastroenterology
IF:22.682
发表时间:2021.8.27
通讯作者:于君
通讯作者单位:香港中文大学
DOI号:10.1053/j.gastro.2021.08.041
实验设计
结果
为了研究HFD在结直肠肿瘤发生中的作用,我们用HFD或CD喂养经AOM处理的常规雄性C57BL/6小鼠,同时进行或不进行定期抗生素鸡尾酒治疗 (图1A)。在取样前进行小鼠结肠镜检查,饲喂HFD的小鼠出现肉眼可见的肿瘤,但饲喂CD或用抗生素治疗的HFD喂养的小鼠没有观察到明显的肿瘤 (图1B)。取样时,饲喂HFD的小鼠的体重和脂肪量均高于饲喂CD的小鼠 (P<0.0001),然而服用抗生素引起肠道微生物群减少的HFD喂养的小鼠,其体重和脂肪量没有影响 (图1C)。饲喂HFD的小鼠的结直肠癌数量和体积也明显大于饲喂CD的小鼠 (均为P<0.05;图1D) 。结肠切片的组织学检查 (图1E) 证实,与饲喂CD的小鼠相比,饲喂HFD的小鼠发生腺癌、高度和低度异长增生的比例更高 (P<0.05;图1E)。饲喂HFD的小鼠结肠切片显示Ki-67阳性细胞明显增多,表明HFD喂养的小鼠细胞增殖增加 (P<0.01;图1F)。然而,饲喂HFD的小鼠肠道微生物群被抗生素消耗后,结肠癌的数量和体积都显著减少 (均为P<0.01;图1D),但饲喂CD的小鼠没有显著减少 (附图1)。结肠切片的组织学检查 (图1E) 证实,服用抗生素的HFD组小鼠的腺癌、高度和低度异常增生的比例低于未服用抗生素的HFD组 (P<0.01;图1E),同时Ki-67阳性细胞显著减少 (P<0.05;图1F)。
为了验证这些发现,我们设置了转基因Apcmin/+小鼠模型(图2A)。与AOM诱导的结直肠肿瘤小鼠模型中的结果一致,HFD显著增加Apcmin/+小鼠的体重和脂肪量(均为P<0.05),并且抗生素治疗后HFD喂养对小鼠的体重和脂肪量没有影响(图2B)。HFD喂养的Apcmin/+小鼠结直肠肿瘤数目显著高于CD喂养的Apcmin/+小鼠(P<0.05;图2C),也表现出肿瘤体积增大的趋势(P=0.07;图2C)。此外,服用抗生素减少肠道微生物群显著降低了HFD喂养的Apcmin/+小鼠的肿瘤数量(P<0.01)和体积(P<0.01;图2C),但在CD喂养的Apcmin/+小鼠中没有(补充图2A和2B)。此外,在HFD喂养的Apcmin/+小鼠的小肠中也观察到肿瘤体积增大,而抗生素诱导肠道微生物群减少显著降低肿瘤体积(补充图2C)。结肠切片的组织学检查(图2D)证实,与CD喂养的小鼠相比,HFD喂养的小鼠发生更大比例的腺癌和高度异常增生(P<0.01;图2D)。HFD喂养的Apcmin/+小鼠与CD喂养的Apcmin/+小鼠相比,Ki-67阳性细胞明显增多(P<0.05;图2E)。而抗生素治疗的HFD喂养的Apcmin/+小鼠显示腺癌、高度和低度异常增生的比例降低(P<0.01;图2D)与无抗生素处理的HFD喂养小鼠相比,病理性Ki-67阳性细胞显著减少(P<0.05;图2E)。这些发现一致表明HFD促进结肠直肠肿瘤发生,并且肠道微生物群可能在介导HFD相关的CRC发展中起重要作用。
图 2. 在 Apcmin/+小鼠模型中,HFD 依赖于肠道微生物群促进 CRC 发展。(A) Apcmin/+小鼠模型饮食和抗生素治疗的实验设计。(B) 第20周结肠镜检查的典型图像。(C)CD喂养、HFD喂养和Abx处理的HFD喂养的Apcmin/+小鼠在处死前的体重和脂肪量。(D) CD喂养、HFD喂养和Abx处理的HFD喂养小鼠处死前的结直肠肿瘤数量和肿瘤体积的代表性图像。(E) HE染色对小鼠结直肠的病理诊断。病理评分按以下标准定量分析: 正常0分; 1为LGD; 2为HGD; 3为癌症。(F) 小鼠结直肠免疫组化染色,定量分析Ki-67指数。AOM,偶氮甲烷; CRC,结直肠癌;CD,控制饮食;HFD,高脂肪饮食;Abx,抗生素;HE,苏木精和伊红; LGD, 低度发育不良; HGD,高度发育不良; IHC,免疫化学。
为了探索肠道微生物失调在HFD相关CRC发展中的潜在作用,我们在AOM模型中对来自CD喂养,HFD喂养和抗生素治疗的HFD喂养小鼠的粪便样品进行了鸟枪宏基因组测序。通过主成分分析(PCA)对CD喂养,HFD喂养和抗生素治疗的HFD喂养的小鼠的微生物群组成进行鉴定(图3A)。与CD喂养的小鼠相比,HFD喂养的小鼠中细菌多样性降低以及细菌丰富度降低(图3A)。在HFD喂养的小鼠中有几种细菌类群存在差异(图3B)。HFD喂养的小鼠中潜在致病细菌种类(包括Alistipes sp. Marseille-P5997和Alistipes sp. 5CPEGH6)的丰度显著高于CD喂养的小鼠(均为P<0.01;图3C);同时,两种保护性细菌,Parabacteroides distasonis(狄氏副拟杆菌,P. distasonis)和Parabacteroides sp. CT06在HFD喂养的小鼠中显著减少(均为P<0.01;图3C)。通过定量PCR验证了与CD喂养的Apcmin/+小鼠相比,在HFD喂养的Apcmin/+小鼠中Alistipes sp. Marseille-P5997和P. distasonis的差异丰度发生典型失调(补充图3)。另外的共培养实验表明P. distasonis抑制细胞生长,Alistipes sp.促进CRC细胞系(Caco-2和HCT116)的细胞生长(图3D),表明HFD诱导的肠道微生物组成的变化可能至少部分有助于CRC的发展。
为了验证肠屏障功能是否在HFD相关的结直肠肿瘤发生中起作用,我们通过测定血清LPS水平来检测HFD对小鼠结肠细胞旁通透性的影响。与用AOM处理的CD喂养的小鼠相比, HFD喂养的小鼠血清LPS浓度升高 (P<0.05;图3E)。与此一致,透射电镜下的肠道屏障结构证实了HFD喂养的小鼠结肠细胞间连接的异常,包括顶端连接复合体和细胞旁间隙的间隙扩大,表明屏障功能受损 (图3F)。此外,HFD喂养的小鼠紧密连接蛋白E-cadherin (一种细胞粘附分子) 和Claudin-3 (紧密连接的关键成分,作为肠道屏障完整性的标志) 的表达显著减少 (均为P<0.01;图3G)。E-cadherin和Claudin-3蛋白的分布主要定位于CD喂养小鼠的结肠上皮细胞膜上,但在HFD喂养的小鼠中它们的表达被分解 (均为P<0.01;图3H)。而服用抗生素的HFD小鼠,其肠道屏障结构得到改善 (图3F),E-cadherin和Claudin-3的蛋白表达恢复 (E-cadherin, P<0.01;Claudin-3,P<0.05) (图3G),E-cadherin和Claudin-3的分布和密度增加 (E-cadherin, P<0.01;Claudin-3,P<0.05) (图3H)。综上所述,这些结果表明HFD至少部分是通过引起肠道微生物失调而导致结肠屏障功能受损的。
图 3. HFD 诱导肠道微生物群失调和肠道屏障功能障碍。(A) AOM 治疗的 CD 喂养、HFD 喂养和 Abx 治疗的 HFD 喂养的小鼠的无监督主成分分析、Shannon多样性和肠道微生物群的物种丰富度。(B) CD 喂养、HFD 喂养和 Abx 处理的 HFD 喂养小鼠肠道微生物组的热图。(C) 与用 AOM 治疗的 CD 喂养或 Abx 治疗的 HFD 喂养小鼠相比,HFD 喂养小鼠中的Alistipes sp. Marseille-P5997,Alistipes sp. 5CPEGH6,P. distasonis 和 Parabacteroides sp. CT06的丰度。(D) Caco-2 和 HCT116 CRC 细胞与 P. distasonis、Alistipes sp、大肠杆菌和空白对照共培养的生长曲线。(E) AOM 模型中 CD 喂养和 HFD 喂养小鼠血清中的 LPS 浓度。(F) 通过 TEM 显示 CD 喂养、HFD 喂养和 Abx 处理的 HFD 喂养小鼠的细胞间连接的代表性图像。(G) 通过蛋白质印迹和定量分析,肠道屏障相关蛋白 E-cadherin 和 Claudin-3 在 CD 喂养、HFD 喂养和 Abx 处理的 HFD 喂养小鼠的结直肠组织中的表达水平。(H) 免疫组化定量分析CD喂养、HFD喂养和Abx喂食HFD喂养小鼠结直肠组织中粘附分子 E-cadherin 和 Claudin-3的分布 *P < 0.05,**P < 0.01, *** P < 0.001。CD,控制饮食;HFD,高脂肪饮食;Abx,抗生素;TEM,透射电子显微镜;IHC,免疫化学。
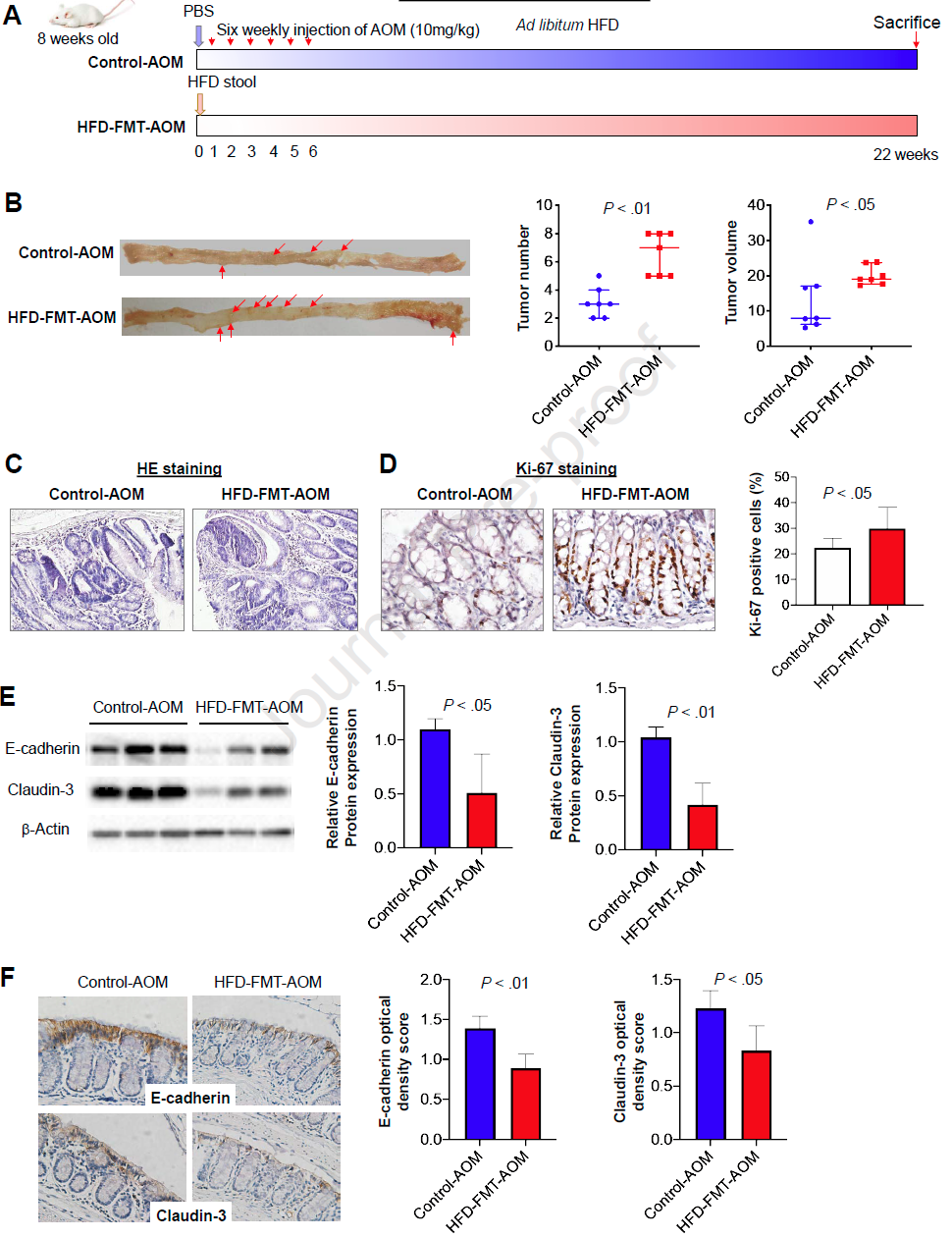
为了进一步验证HFD调节的微生物群对结直肠肿瘤发生的影响,我们将HFD喂养的小鼠的粪便样本转移到用AOM处理的无菌小鼠 (图4A)。粪便微生物群移植没有改变AOM处理的无菌小鼠的体重和脂肪量 (补充图4A)。用HFD喂养的小鼠的粪便样品灌胃的无菌小鼠与灌胃PBS的AOM处理的无菌小鼠相比 (图4B), 另枝菌属显著富集 (P<0.001,补充图4B),结直肠肿瘤的数量增多 (P<0.01) 以及肿瘤的体积增大 (P<0.05)。结直肠肿瘤经组织学证实为低度异常增生 (图4C)。与此相一致的是,AOM处理的HFD喂养的小鼠(HFD-FMT-AOM)结肠组织中Ki-67阳性细胞较对照组AOM处理的小鼠明显增多 (P<0.05;图4D)。此外,与对照组小鼠相比,灌胃HFD小鼠粪便标本的小鼠结直肠组织中E-cadherin和Claudin-3的表达明显减少 (E-cadherin和Claudin-3分别为P<0.05和P<0.01;图4E)。同样,E-cadherin和Claudin-3蛋白在HFD-FMT-AOM小鼠中被分解,但在对照AOM小鼠的结肠上皮细胞膜上易表达 (E-cadherin和Claudin-3分别为P<0.01和P<0.05;图4F)。综上所述,这些结果表明,HFD调节的肠道微生物群通过损害肠道屏障功能促进了结直肠肿瘤的发生。
为了揭示由HFD诱发的代谢变化,我们对用AOM处理的CD喂养的小鼠,HFD喂养的小鼠和抗生素治疗的HFD喂养的小鼠的粪便样品进行代谢谱分析。根据无监督PCA分析和PLS判别监督分析(PLS-DA),发现膳食脂肪摄入和抗生素治疗的肠道代谢物显著不同(图5A)。与CD喂养的小鼠相比,在HFD喂养的小鼠中鉴定出差异的代谢物(图5B,补充表4)。其中,包括溶血磷脂酰胆碱(LPC)和溶血磷脂酸(LPA)(LPC的下游代谢物)在内的甘油磷脂都是HFD喂养小鼠中上调最多的异常代谢物(图5B,补充图5)。与CD喂养的小鼠相比,HFD喂养的小鼠中上调的代谢物富含不同的代谢组学信号传导途径。在这些途径中,甘油磷脂代谢是喂养HFD的小鼠中改变的主要途径(图5C)。另一方面,与HFD喂养的小鼠相比,抗生素治疗的HFD喂养小鼠中肠道微生物群的减少显著恢复了已改变的代谢物(图5D,补充表5),LPA和LPC水平降低(图5D,补充图5)。同样地,肠道微生物群的消耗减少了包括甘油磷脂代谢在内的途径(图5E)。
我们通过综合分析确定了HFD喂养的小鼠肠道微生物变化和代谢产物之间的潜在联系。发现Alistipes sp. Marseille-P5997, Alistipes sp. 5CPEGH6, Bifidobacterium animalis(动物双岐杆菌) 以及 Alistipes shahii的富集与LPA、LPC均呈正相关 (图5F)。而Parabacteroides sp. CT06 和P. distasonis与LPA和LPC均呈负相关 (图5F)。这些结果表明,肠道微生物群及其相关的代谢物的改变都有助于HFD相关的结直肠肿瘤的发生。
用差异和非差异代谢物处理两种CRC细胞系(Caco-2和HCT116)和一种正常结直肠上皮细胞系(NCM460)去探索HFD改变的代谢物在CRC发展中的潜在作用。与CD喂养的小鼠相比,HFD喂养的小鼠中的去甲二氢愈创木酸(NDGA)和神经酸显著减少(补充图6A)。肌苷是HFD喂养的小鼠和CD喂养的小鼠之间的非差异代谢物之一,用作阴性对照。共培养实验表明,NDGA通过抑制细胞生长,显著抑制CRC细胞系Caco-2和HCT116的细胞增殖,但不抑制正常上皮细胞系NCM460的细胞增殖(均为P<0.01;图6A)和集落形成(均为P<0.01;图6B)。同样,神经酸也抑制Caco-2和HCT116细胞生长(均为P<0.01;补充图6B)和集落形成(均为P<0.05)(补充图6C)。此外,我们分别用NDGA和神经酸处理患者来源的CRC类器官,发现NDGA和神经酸均可显著抑制CRC类器官生长(均为P<0.01,补充图6D)。然后,我们关注HFD喂养后富集的代谢物LPA的致癌作用。我们发现LPA能显著促进三种结直肠细胞系的增殖(均为P<0.01;图6C)。细胞周期分析显示,与空白对照相比,LPA处理加速了HCT116中从G1期到S期的细胞周期进程,以及Caco-2细胞中从G1期到S期和G2期的细胞周期进程(图6D)。同样,经LPA处理后的CRC细胞系中发现增殖细胞核抗原(PCNA),细胞周期蛋白D1和已下调的P27蛋白表达上调(图6E)。另外,我们使用或不使用LPA处理患者来源的CRC类器官以进一步证实LPA的致癌作用,并证明LPA显著促进CRC类器官的生长和增殖(图6F)。此外,我们还检查了LPA是否会影响上皮屏障功能。与其空白对照相比,在LPA处理的HCT116,Caco-2和NCM460细胞中观察到上皮细胞钙粘蛋白和闭合蛋白表达显著降低,推断LPA可能损害屏障功能(图6G)。总之,这些结果表明HFD至少部分通过提高致癌代谢物LPA和减少潜在的有益代谢物NDGA和神经酸来促进CRC发展。
 图
6. HFD改变的代谢物有助于细胞增殖和细胞连接损伤。
(A)用NDGA和肌苷(作为阴性对照)处理的CRC细胞系Caco-2,HCT116和正常结肠细胞系NCM460的细胞生长曲线。
(B)用NDGA和肌苷(作为阴性对照)处理的CRC细胞系中集落形成的代表性图像和定量分析。
(C)用LPA和肌苷(作为阴性对照)处理的CRC细胞系Caco-2,HCT116和正常结肠细胞系NCM460的细胞生长曲线。
(D)用或不用LPA处理的HCT 116细胞和Caco-2细胞用碘化丙啶(PI)染色并通过流式细胞术分析。
(E)用LPA和肌苷处理的HCT 116和Caco-2细胞中细胞增殖和细胞周期相关蛋白PCNA,细胞周期蛋白D1和P27的表达水平。
(F)用LPA和肌苷处理的CRC类器官的代表性图像和类器官活力测定。
(G)肠屏障功能相关蛋白E-cadherin和Occludin在HCT116,Caco-2和NCM460细胞系中的表达水平。
LPA,溶血磷脂酸;
PCNA,增殖细胞核抗原;
CRC,结直肠癌。
图
6. HFD改变的代谢物有助于细胞增殖和细胞连接损伤。
(A)用NDGA和肌苷(作为阴性对照)处理的CRC细胞系Caco-2,HCT116和正常结肠细胞系NCM460的细胞生长曲线。
(B)用NDGA和肌苷(作为阴性对照)处理的CRC细胞系中集落形成的代表性图像和定量分析。
(C)用LPA和肌苷(作为阴性对照)处理的CRC细胞系Caco-2,HCT116和正常结肠细胞系NCM460的细胞生长曲线。
(D)用或不用LPA处理的HCT 116细胞和Caco-2细胞用碘化丙啶(PI)染色并通过流式细胞术分析。
(E)用LPA和肌苷处理的HCT 116和Caco-2细胞中细胞增殖和细胞周期相关蛋白PCNA,细胞周期蛋白D1和P27的表达水平。
(F)用LPA和肌苷处理的CRC类器官的代表性图像和类器官活力测定。
(G)肠屏障功能相关蛋白E-cadherin和Occludin在HCT116,Caco-2和NCM460细胞系中的表达水平。
LPA,溶血磷脂酸;
PCNA,增殖细胞核抗原;
CRC,结直肠癌。
为了评估HFD调节的肠道微生物群和代谢产物对肠道细胞增殖和致瘤性转化的直接影响,用HFD喂养的小鼠 (HFD-FMT) 和CD喂养的小鼠 (CD-FMT) 的粪便灌胃无菌小鼠。所有这些无菌小鼠都喂以正常的食物,在实验过程中没有任何干扰(图7A)。与CD-FMT小鼠相比,HFD-FMT小鼠的体重和脂肪量没有差异 (补充图7)。与CD-FMT小鼠相比,HFD-FMT小鼠粪便可促进结肠细胞显著增殖,Ki-67阳性细胞增多 (P<0.01;图7B),PCNA蛋白表达增多 (P<0.05;图7C)。为了从机制上了解HFD调控的肠道微生物群/代谢物如何促进结直肠增殖,利用Mouse Cancer Pathway Finder PCR Array检测了HFD-FMT小鼠结直肠组织的基因表达谱,发现HFD-FMT小鼠与CD-FMT小鼠相比,癌基因上调和抑癌基因下调,这些基因主要参与调控细胞增殖 (CCND2,McM2,Mki67)、上皮细胞-间充质转化 (GSC)、缺氧信号转导 (EPO) 的表达(图7D)。
我们证实了与CD-FMT小鼠相比,HFD-FMT小鼠的血清LPS水平显著升高(P<0.05),E-cadherin和Claudin-3的结直肠蛋白显著降低 (均为P<0.01;图7E), HFD相关肠道微生物群在HFD相关性结直肠癌的发生发展中起到了肠道屏障功能障碍的作用。相应地,HFD-FMT小鼠与CD-FMT小鼠相比,E-cadherin和Claudin-3的蛋白表达和密度均显著降低 (均为P<0.01;图7F)。我们评估了受体无菌小鼠肠道微生物群的组成,发现HFD-FMT小鼠与CD-FMT小鼠肠道微生物群显著不同 (图7G),包括潜在致病菌Alistipes sp. 5CPEGH6 和 Alistipes sp. Marseille P5997的富集 (均为P<0.05;图7H)。综上所述,这些数据表明HFD相关的肠道微生物群/代谢物直接促进结直肠细胞增殖并损害肠道屏障功能。
讨论
在本研究中,我们证明了高脂肪含量的饮食促进了不同小鼠模型的结直肠肿瘤的发生。有研究表明西方饮食与人类结直肠癌风险的增加有关,这证明了HFD在结直肠肿瘤发生中的促肿瘤作用。食用高脂饮食通常会导致肥胖,这会使男性患结肠癌的风险增加30%-70%。多项证据表明,肥胖相关的脂肪细胞因子、胆汁酸失衡和肠道微生物群失调均有助于结直肠癌的发生。虽然在AOM处理模型和Apcmin/+模型中,HFD喂养的小鼠的体重、脂肪量和肿瘤负荷都有所增加,但抗生素减少肠道微生物群降低了肿瘤负荷,但不改变体重或脂肪量。无菌模型进一步证实了这一点,在无菌模型中,转移了HFD相关肠道微生物群的小鼠存在更大的肿瘤负担以及细胞增殖加快,但没有显著的体重变化。这些发现共同表明HFD/肥胖相关的肠道微生物群在结直肠肿瘤的发生中起着重要作用。
接下来,我们发现HFD喂养的小鼠表现出不同的微生物群组成,因此暗示了肠道微生物群失调的发生。Alistipes sp. Marseille-P5997 和 Alistipes sp. 5CPEGH6在HFD喂养的小鼠中显著富集。特别是,在食用HFD的人类中也发现Alistipes属的富集。Alistipes与结直肠肿瘤负担呈正相关。此外,将我们的小鼠模型的微生物群结果与结直肠癌患者和对照组的结果相比较,发现Alistipes在小鼠模型和人类结直肠癌患者中均有富集,这表明HFD诱导的结直肠癌小鼠模型肠道微生物群改变与结直肠癌患者的特征有一定的相似之处。同样,我们的共培养实验表明,Alistipe sp.可以促进结直肠癌细胞生长 (图3D)。此外,据报道Alistipes finegoldii能够通过IL-6/STAT3途径促进了CRC的形成。综上所述,Alistipes在结直肠肿瘤的发生中作为潜在的病原体。另一方面,两种益生菌P. disasonis和Parabacteroides sp. CT06在HFD喂养的小鼠体内减少。本研究证实了P. disasonis的保护作用,它能抑制体外培养的结直肠癌细胞的生长。此外,已有的研究证实P. disasonis通过阻断Toll样受体4信号通路和Akt激活来减少体内结肠肿瘤的形成。此外,P. disasonis还参与维持肠上皮屏障功能,以抑制结直肠癌的发展。我们进一步证实,HFD调节的肠道微生物群可以促进AOM处理的无菌小鼠的结直肠癌的发生,这表明用HFD喂养的小鼠的粪便样本灌胃的无菌小鼠结肠癌的形成明显增加,并伴随着肠道屏障功能的受损。这些结果证实HFD所致肠道微生物失调,致癌病原菌富集,保护菌减少,在HFD相关结直肠癌的发生发展中起重要作用。
然后,我们探讨了由HFD诱发改变的上皮屏障功能及其相关的肠道微生物群变化。在HFD喂养的小鼠中观察到血清LPS水平升高。此外,在HFD喂养的小鼠的结直肠中顶端连接复合物和细胞间隙的空间变大以及肠屏障相关标志蛋白质: E-cadherin和Claudin-3的表达降低,表明肠屏障功能受损。然而,我们发现向HFD喂养的小鼠服用抗生素以减少肠道微生物群,恢复了受损的肠道屏障功能,推断肠道微生物生态失调参与了HFD相关的肠道屏障功能障碍。一些饮食方案可以推动微生物组的转变,而这反过来又会以各种方式导致屏障的破坏。粘膜屏障的破坏可能导致共生微生物(包括病原体)无限制的通过固有层甚至血液,使其更容易影响CRC的发展。此外,在结直肠粘膜的组织病理学变化之前或之后观察到CRC患者中LPS水平增加和动物模型中结直肠通透性增加,表明肠道屏障受损有助于CRC发展。总而言之,这些结果表明HFD诱发的结直肠肿瘤的发生与肠道屏障功能受损有关,其机制可能与肠道生物失调有关。
随后,我们评估了HFD相关的肠道微生物失调在HFD相关的结直肠肿瘤发生中的作用。在两种常规模型中,服用抗生素减少肠道微生物群,显著降低了HFD相关肿瘤的发生,表明HFD相关的小鼠结直肠肿瘤发生与完整微生物群的存在有关。利用无菌动物模型,我们进一步验证了HFD调控的肠道微生物群可以直接促进AOM处理的无菌小鼠结直肠肿瘤的发生。我们的结果进一步得到了两种肠道肿瘤小鼠模型K-rasG12Dint小鼠和Apcmin/+小鼠的研究结果的支持,这两种小鼠模型在遗传上易患肠道肿瘤。与没有粪便微生物组移植的K-rasG12Dint小鼠相比,将HFD喂养的肠道肿瘤小鼠的粪便样本转移到经过充分抗生素治疗的健康成年K-rasG12Dint小鼠,可以诱导小肠肿瘤的发展和病理评分的增加。在接受相同粪便微生物群转移的Apcmin/+小鼠中也发现了类似的情况。此外,抗生素治疗完全阻断了HFD诱导的K-rasG12Dint小鼠的肿瘤进展,这与我们在AOM处理的Apcmin/+小鼠中的结果一致,表明微生物群的明显变化在加剧肠道癌变中起着关键作用。综上所述,我们的发现表明肠道微生物群在调节HFD相关的结直肠肿瘤的发生中是必不可少的。
除了肠道菌群失调之外,由膳食食物和共生细菌产生的小分子代谢物也可能有助于CRC的发展。肠道微生物相关代谢物可以激活和拮抗其同源受体,以减少或加剧肠道肿瘤的发展。因此,我们揭示了肠道代谢物改变对HFD相关结直肠肿瘤发生的作用。与CD喂养的小鼠相比,HFD喂养的小鼠的代谢组学特征发生了明显的变化。NDGA是一种经典的酚木脂素,在HFD喂养的小鼠中减少。我们证明NDGA可以显著抑制CRC细胞生长和集落形成。动物研究进一步支持了这一观点,有研究报道NDGA可预防肺癌发生并抑制异种移植模型中非小细胞肺癌的生长,这表明NDGA可能是癌症治疗中潜在的治疗补充剂。除NDGA外,还发现神经酸在HFD喂养的小鼠中减少。我们发现神经酸抑制CRC细胞增殖和集落形成。据报道,在肥胖小鼠模型中神经酸减少,补充神经酸可以减轻HFD喂养的小鼠的体重和脂肪量,并改善代谢参数,这表明神经酸是预防肥胖和CRC的潜在补充剂。
更重要的是,我们发现在具有完整肠道微生物群的HFD喂养的小鼠中LPA和LPC水平升高,但在肠道微生物组减少的小鼠中没有,并促进小鼠CRC发展。此外,证明了LPA的促肿瘤发生功能,其中LPA促进CRC细胞系和患者衍生的CRC类器官中的细胞增殖并加速细胞周期。升高的LPA还损害CRC细胞系中的上皮细胞连接。LPA和LPC都可能是磷脂酶A2组1B(PLA2g1b)被肠道微生物群操纵产生的生物活性脂肪分解产物。LPA的致癌特性已在不同的癌症中得到证实。最近的研究表明,autotaxin-LPA轴促进胰腺导管腺癌细胞增殖,阻断autotaxin-LPA轴可以抑制体内胰腺癌生长,表明LPA可能是癌症治疗的药物靶标。总而言之,我们的研究结果表明,HFD及其相关的肠道微生物群失调,导致致癌LPA水平升高,促进细胞增殖并损害肠道屏障,从而促进CRC的发展。
最后,我们探讨了HFD调节的肠道微生物群/代谢产物在正常饮食条件下对无菌小鼠健康结肠粘膜的直接影响。在正常饮食条件下,接受HFD小鼠粪便的无菌小鼠细胞显著增殖,表明HFD调节的肠道微生物群/代谢产物在促进细胞增殖方面起着直接作用。从机制上,我们揭示了HFD调控的肠道微生物群/代谢物可以上调癌基因的表达,这些基因主要参与调控细胞增殖和上皮向间质的转变。肠道微生物群具有调节遗传或表观遗传表达的能力。以往的研究表明,肠道微生物群可通过WNT信号通路的过度激活影响突变型p53促进肠道肿瘤形成。此外,将结直肠癌患者的粪便移植到无菌小鼠身上,与细胞增殖、凋亡、血管生成和转移相关的基因发生上调,这支持了我们在这项研究中的结论。此外,与CD-FMT无菌小鼠相比,HFD-FMT无菌小鼠的肠道屏障相关蛋白表达减少,表明HFD调节的肠道微生物群在结直肠癌发病前的肠道屏障功能受阻中起直接作用。这与结肠通透性增高可能早于结肠粘膜的组织病理学改变的报道一致。HFD-FMT小鼠和CD-FMT小鼠在肠道菌群组成上存在显著差异,提示肠道菌群失调可能是导致肠道屏障功能障碍的触发因素之一。这些结果进一步表明,摄入过量脂肪引起肠道微生物群/代谢产物的改变,加速了细胞增殖和肠道屏障功能障碍,导致结直肠癌的发生。
总之,我们的研究发现,HFD通过诱导小鼠肠道微生物失调、代谢改变和肠道上皮屏障功能受损来推动结直肠癌的发生 (补充图8)。HFD调控的肠道微生物群可能通过上调癌基因表达,损害肠道屏障功能,从而促进结直肠癌的发生。调节肠道微生物群和代谢产物可能是预防和治疗HFD相关结直肠癌的潜在治疗策略。
不感兴趣
看过了
取消

人点赞

人收藏

打赏
不感兴趣
看过了
取消
©2012-2023 北京华媒康讯信息技术股份有限公司 All Rights Reserved. 注册地址:北京 联系电话:010-82736610
广播电视节目制作经营许可证 —(京)字第 17437号 京海食药监械经营备20200522号
京ICP备12011723号 京ICP证150092号
 京公网安备 11010802020745号
工商备案公示信息
互联网药品信息服务资格证书((京)-非经营性-2020-0015)
京公网安备 11010802020745号
工商备案公示信息
互联网药品信息服务资格证书((京)-非经营性-2020-0015)






您已认证成功,可享专属会员优惠,买1年送3个月!
开通会员,资料、课程、直播、报告等海量内容免费看!

打赏金额
认可我就打赏我~
1元 5元 10元 20元 50元 其它
打赏作者
认可我就打赏我~
扫描二维码
立即打赏给Ta吧!
温馨提示:仅支持微信支付!
已收到您的咨询诉求 我们会尽快联系您






































 010-82736610
010-82736610
 股票代码: 872612
股票代码: 872612




