
这项研究表明手术应激可通过ERK5 /整合素GPIIb / IIIa信号激活血小板,最终导致血小板-CTC聚集增加。

外科切除是治疗绝大多数实体瘤的主要手段,但是手术自身也会增加肿瘤复发和转移的风险。组织创伤会局部和系统地激活先天免疫系统,从而引起炎症反应。血小板和中性粒细胞是早期先天免疫反应中促进组织愈合的两个关键因素,但它们也可能有助于癌细胞的扩散和远处转移。
2021年3月俄亥俄州立大学医学中心Allan Tsung教授和Hai Huang教授在《Cancer Research》上发表了题为《Platelet TLR4-ERK5 axis facilitates NETs-mediated capturing of circulating tumor cells and distant metastasis after surgical stress》的论著。论文细致地描述了血小板TLR4-ERK5轴在促进中性粒细胞胞外诱捕网(NETs)介导的循环肿瘤细胞捕获中发挥的作用,并提出靶向血小板活化通路TLR4/ERK5/integrin GPPIIb/IIIa具有防止术后NETs介导的肿瘤远处转移的治疗前景。
这篇文章报道了外科手术应激激活的血小板会促进血小板-肿瘤细胞聚集体的形成,从而促进它们被中性粒细胞胞外诱捕网(NETs)捕获和随后的远处转移。小鼠肝脏局部缺血/再灌注(I /R,以下用肝I/R代替)损伤局部手术应激模型显示,I / R促进NETs对聚集的循环肿瘤细胞(circulating tumor cells, CTCs)的捕获并最终转移至肺部,而去除掉血小板时上述现象消失。肝I / R也增加了NETs在肺微脉管系统中的沉积,然而去除掉血小板对NETs沉积没有影响。
TLR4对于ERK5-GPIIb / IIIa整合素依赖性方式的血小板活化和血小板-肿瘤细胞聚集体形成至关重要。阻断血小板活化或敲除TLR4可导致NETs对CTC没有捕获作用,最终可保护小鼠免受肝I / R诱导的转移影响。上述结果首次发现血小板和中性粒细胞在急性炎症的情况下有助于肿瘤转移,而靶向血小板与NETs之间的相互作用具有防止术后肿瘤远处转移的治疗前景。
从杰克逊实验室购买雄性C57BL / 6小鼠(8至12周)。在匹兹堡大学的Timory Billiar博士的实验室中制作并繁殖了TLR4基因敲除(TLR4-/-),TLR4 flox(TLR4loxp / loxp)和血小板特异性TLR4-KO(PF4-TLR4-/-)小鼠。在我们的实验室饲养了IV型肽基精氨酸脱亚氨酶(PAD4-/-)小鼠。鼠结肠癌MC38细胞获自Michael Lotze博士。CT26鼠结肠直肠癌细胞购自ATCC。两种细胞系均使用基因组图谱进行了鉴定(IDEXX Radil Cell Check)。使用MycoFluor支原体检测试剂盒对细胞进行常规测试以确保其不含支原体。
所有实验均符合动物伦理。为了建立实验性肺转移模型,我们通过尾静脉给小鼠静脉注射混悬于100 µl PBS中的MC38细胞或CT26细胞。使肿瘤细胞循环10分钟。接着我们对小鼠进行肝缺血/再灌注造模,假手术组不进行肝缺血造模。
肺标本用4%多聚甲醛的磷酸盐缓冲盐水(PBS)固定,并包埋在O.C.T.化合物,并切成8微米的厚度。将肺切片与针对Ly6G,Cit-H3,CD41和DAPI的抗体孵育。通过Z平面获得X和Y的大面积图像,并使用尼康A1共焦显微镜(显示为Z的平均强度投影。使用NIS Elements Software(Nikon)进行定量。
机械分解肺组织,然后在DMEM中的胶原酶IV(5 114 mg / ml)和DNase(100 U / ml)溶液中酶消化37℃。然后将部分消化的组织通过70 µm细胞过滤器过滤。使用LSRFortessa流式细胞仪获取流式细胞仪数据,并通过Flowjo软件进行分析。
通过使用流式细胞仪和Alexa Fluor 488标记的血纤蛋白原(与活化的GPIIb / IIIa特异性结合)来确定血小板GPIIb / IIIa的活化程度。将经过肝脏IR或假手术的小鼠的富含血小板的血浆(PRP)与15 µg / ml Alexa Fluor 488标记的纤维蛋白原在37°C孵育15分钟,用2%多聚甲醛固定20分钟,并进行流式分析细胞计数。
将血小板从经历了1.5小时的部分肝局部缺血,然后再进行3小时的再灌注或假手术的小鼠中分离出来,在用血球计数器手动计数后,将血小板的密度调整为2×107/ ml。在37°C用5μMA23187刺激源自骨髓的中性粒细胞(1×106)长达3小时以诱导NETs的形成。为了与NETs一起温育,将1×105 MC38细胞与2×107血小板预温育3小时,然后在37°C的摇床温箱中加入终体积为1.5 ml的装有NETs的试管中6小时。将细胞在染色溶液(含有1%BSA和2mM EDTA的PBS)中与特定抗体在适当的稀释度下孵育30分钟。然后将细胞用PBS洗涤,并立即通过流式细胞术进行分析。
使用来自肺组织或洗涤过的血小板的全细胞裂解液进行了蛋白质印迹分析。我们采用了以下抗体:Cit-H3(1:800 Abcam),ERK5(1:800 CST),pERK5(1:800 CST),p70S6K(1:800 CST),Rac1(1:800 Abcam),actin作为对照。
为了从小鼠中瞬时清除血小板,在I / R前12小时腹膜内注射30μg抗CD41抗体(MWReg30,Abcam)。而在注射肿瘤和肝I / R造模之前,将从野生型或TLR4-/-小鼠中新鲜分离的血小板以每只小鼠2×107的细胞剂量静脉内注射到血小板已被去除的小鼠中。
图中显示的数据为平均值±SEM(平均值的标准误差)。使用单因素方差分析与事后Tuker(HSD)分析和Student's t检验进行组比较。使用Spearman相关系数检查MPO-DNA水平与肿瘤细胞数量之间的相关性。P <0.05的值被认为具有统计学意义。GraphPad Prism 7软件用于所有统计分析和生成图形。
为了确定手术后的局部炎症反应是否促进CTCs在潜在转移部位的滞留,我们使用了高度标准化的部分(70%)肝I / R小鼠手术应激模型。通过尾静脉将1×106羧基荧光素琥珀酰亚胺酯(CFSE)标记的MC38鼠结肠癌细胞注入小鼠全身循环,并建立肝I / R模型或假手术。
注射后第21天,我们发现,与假手术组相比,肝I / R组小鼠在肺表面的转移灶数量高约高三倍(P <0.001)(图1A),这表明外科手术应激在肿瘤转移中的重要作用。对苏木精和曙红染色的肺组织切片上转移灶的覆盖范围的定量分析结果显示:肝I / R组的每肺转移灶面积为53.5%,而假手术组仅为4.6%(P <0.001)(图1B)。
为了深入了解肝I / R如何引起远处转移,我们计数了在尾静脉注射后不同时间点肝I / R的小鼠肺中的肿瘤细胞。一般情况下,由于血液动力学的剪切力和免疫系统的攻击,只有少数循环中的肿瘤细胞能够存活并外渗以在远处形成转移。有趣的是,在注射肿瘤细胞后2小时内,在肝I / R组和假手术组小鼠的肺中CFSE标记的MC38细胞数量的无显著统计学差异(图1C)。但是,在稍后的时间点(6小时,12小时和24小时),肝I / R导致肿瘤细胞显著滞留(图1C,1D和1E)。同时,将不同数量的CFSE标记的MC38细胞注射入小鼠体循环,然后进行肝I / R或假手术。该结果与先前的结果一致:在接受肝I / R的小鼠肺部滞留了更多的肿瘤细胞(图1F)。为了进一步验证其他肿瘤细胞类型在其中的作用,我们使用CT26鼠结肠直肠癌细胞重复了以上实验。结果表明,肝I / R导致小鼠肺中CT26肿瘤细胞的显著滞留(补充图1A)。这些数据表明,由肝脏I / R引起的外科手术应激并不会增加肺毛细血管中CTCs的数量,而是促进循环中CTCs的更好存活以及随后在转移过程中的定植。

肿瘤细胞进入循环后的最初几个小时内,动态的宿主-肿瘤细胞相互作用决定了远处转移是否成功。血小板可以在几分钟内在肿瘤细胞周围形成血小板-肿瘤聚集体,从而导致肿瘤细胞在血管内存活。为了确定由手术应激下激活的血小板是否对肿瘤细胞具有更高的亲和力,我们将MC38细胞与从接受肝I / R或假手术的小鼠分离的血小板共培养。与对照组假手术小鼠(占总肿瘤数量的21.8%)相比,肝I / R小鼠(占总肿瘤数量的48.6%)形成血小板-肿瘤聚集体的肿瘤细胞百分比显著增加(P <0.01)(图2A和2B)。此外,当注射时间从1小时延长至3小时(占肿瘤总数的61.5%)时,MC38细胞的血小板-肿瘤聚集进一步增加(P <0.05)(图2A和2B)。这些结果并不是MC38癌细胞所独有的,肝I / R还会促进血小板与CT26细胞的聚集(附图1B)。接下来,我们试图通过体内实验进行验证。如图2C所示,免疫荧光染色结果表明,与假手术小鼠相比,在接受肝I / R的小鼠肺部,被血小板包围的CFSE标记的MC38细胞(CD41阳性)的数量明显更高。值得注意的是,与对照抗体治疗相比,在肿瘤细胞注射和肝I / R之前,腹膜内注射消耗血小板的抗体(anti-CD41)消耗血小板可导致肺中滞留的肿瘤细胞数量显著减少(P < 0.0001)(图2D和2E)。这些研究数据表明,肝I / R后系统性血小板-肿瘤细胞聚集体的形成可能有助于癌细胞的捕获和随后在肺中的远处转移。
前期我们发现肝脏中的嗜中性粒细胞为响应肝I / R释放了嗜中性粒细胞胞外陷阱(NETs),而这些NETs促进了肝脏内肿瘤的进展。在这项研究中我们则观察到了肝I / R的全身作用。因此,我们假设肝脏炎症也可能触发远端部位(包括肺)的NETs形成,以促进CTCs的捕获。为了确定肝I / R后肺组织内的NETs形成,我们使用了免疫荧光染色来检测测NETs。在肝I / R小鼠模型中的肺组织内我们可以观察到许多Ly6G阳性细胞(鼠中性粒细胞的表面标志物)与细胞外组蛋白和网状DNA的共定位,表明NETs的形成(图3A)。此外,在小鼠肝I / R 建立后的6、12和24小时能检测到肺组织中组蛋白H3(Cit-H3)的瓜氨酸化(一种NETs的标志物)表达增加(图3B)。循环髓过氧化物酶(MPO)-DNA复合物水平(NETs形成的另一种特异性标志物)与再灌注后6小时被困在肺微脉管系统中的转移性肿瘤细胞数量呈正相关(Spearman系数0.969,P <0.0001)(图3C)。用DNase I破坏现有的NETs或通过敲除PAD4抑制NETosis后,肝I / R小鼠和假手术的小鼠之间肺微脉管系统中CTCs数量没有明显差异(图3D)。这些结果表明肝I / R诱导的炎症会导致肺中NETs的形成,并与CTCs在肺微脉管系统中的滞留有关。接着我们继续探究活化的血小板是否会影响肝I / R期间NETs的形成。我们在手术前用抗血小板抗体处理小鼠。我们发现在抗CD41抗体和对照组抗体治疗的小鼠肺中Cit-H3和MPO-DNA复合物水平的无明显差异,这表明在手术应激下NETs的形成与血小板无关(图3E和3F)。
接着我们检测活化的血小板是否对NETs的活性(如捕获CTCs)有影响。我们用钙离子载体A23187刺激髓源性中性粒细胞以形成NETs,然后在肝I / R或假手术小鼠组来源的血小板存在的条件下与MC38细胞共培养。我们通过流式细胞术探究Ly6G阳性细胞与MC38细胞的相互作用,以代表NETs介导的肿瘤细胞捕获。与在对照组中被捕获的肿瘤细胞(无血小板,9.03%)相比,假手术小鼠组中有血小板存在的情况下NETs捕获了更多的肿瘤细胞(17.33%;P> 0.05)(图4A)。令人惊讶的是,从肝I / R小鼠中分离出的血小板导致NETs捕获了很高比例的肿瘤细胞(67.82%)(P <0.001)(图4A)。与上述结果一致的是CT26肿瘤细胞与肝I / R模型来源的血小板共培养也促进了NETs介导的肿瘤捕获。
为在实验室中模拟体内循环及其内环境,我们对动态流动条件进行了模拟,然后评估了在有或没有血小板存在的条件下NETs与肿瘤细胞之间的相互作用。我们采用平行流动室以模拟肺血流状况,即采用上述装置在单层中性粒细胞上灌注MC38细胞或与血小板预孵育的MC38细胞。与我们之前的研究结果一致的是:在灌注10分钟后,经从肝I / R小鼠分离的血小板进行预孵育的NETs会捕获大量的肿瘤细胞(P <0.001)(图4B和补充材料里的影像资料)。相反,用假手术组小鼠来源的血小板对MC38细胞进行预处理对NETs捕获MC38细胞没有影响(图4C)。这与在静态系统下获得的结果一致(图4D)。
NETs对血小板-肿瘤细胞聚集体捕获能力增强意味着血小板可以刺激肿瘤细胞向NETs迁移。我们试图在体外直接通过transwell迁移方法进行验证。NETs在transwell下室中用A23187预先诱导。然后将无FBS培养基的MC38细胞接种到上室中。结果显示:与对照组相比(无血小板,每视野185个),在假手术组来源血小板存在的情况下有更多的MC38细胞通过transwell孔迁移(每视野219个;P <0.05)。此外,在肝I / R小鼠组来源血小板存在的情况下,MC38细胞的迁移能力进一步提高(435 /每视野;P <0.001)(图4E)。综上所述,这些结果表明NETs易于捕获血小板-CTCs聚集体。

接着我们将进一步探究手术应激下的血小板激活机制。活性氧(ROS)是I / R损伤的标志。在心肌梗塞中,活性氧已被证明可通过细胞外调节蛋白激酶5(ERK5)引起血小板活化。因此,我们评估了ERK5在肝I / R激活血小板过程中的作用。与假手术组相比,肝I / R小鼠组来源的血小板中ERK5磷酸化位点Thr 218及Thr 220处磷酸化的表达升高,并且pERK5 / ERK5的比例显著升高(图5A)。此外,肝I / R组血小板中下游通路70 kDa核糖体S6激酶(P70S6K)和Ras相关的C3肉毒杆菌毒素底物1(Rac1)的表达也显著升高,而上述成分是整合素激活所需的两个不可或缺的信号转导成分。
已知活化的血小板可通过多种细胞表面分子与肿瘤细胞相互作用。整联蛋白(主要是GPIIb / IIIa)和P-selection是两种常见的结合肿瘤细胞的血小板表面蛋白。我们用Alexa Fluor 488标记的纤维蛋白原(特异性结合激活的GPIIb / IIIa)来探究整联蛋白的状态。与假手术组小鼠相比,肝I / R小鼠中血小板GPIIb / IIIa的活化更为明显(2092 MFI [1126-2976]与811 MFI [693-984],P <0.01)。然而,在实验组小鼠和对照组小鼠的血小板P-selection蛋白表达的无显著性差异。
我们进一步探究了ERK5信号通路的抑制剂XMD8-92是否会降低肝I / R诱导的GPIIb / IIIa激活,血小板-肿瘤聚集和CTC的肺转移。如预期的那样,XMD8-92显著降低了肝I / R血小板中ERK5的磷酸化。用XMD8-92处理后,P70S6K / RAC1的表达和GPIIb / IIIa的激活降低(图5B和5C)。XMD8-92预处理阻断了肝I / R诱导的血小板肿瘤聚集体的形成(图5D)。另外,与对照组小鼠相比,XMD8-92预处理能降低90%的CTCs肺转移(P <0.001)(图5E和5F)。
为了研究ERK5信号传导是否抑制NETs的形成,我们分别测量MPO-DNA复合物和肺Cit-H3的表达。与我们前述结果一致,肝I / R模型中XMD8-92治疗组与对照组相比血清中循环MPO-DNA复合物水平以及肺中Cit-H3表达水平未见明显差异(图5G和5H)。这进一步支持了我们的发现,即在手术应激下血小板可能不直接参与NETs的形成,而是在后期参与CTCs捕获。综上所述,血小板ERK5-GPIIb / IIIa信号通路在肝I / R期间血小板活化和血小板-肿瘤细胞聚集中起着至关重要的作用。
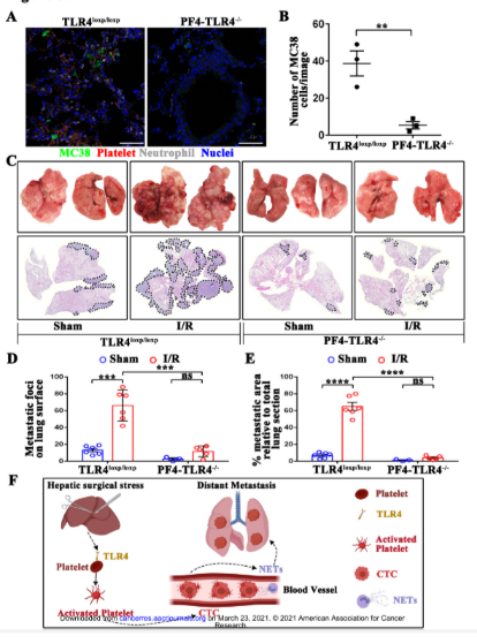
目前外科手术应激期间导致ERK5激活的信号传导途径尚不清楚。血小板表面的Toll样受体具有识别抗原的能力。研究报道血小板TLR4是失血性休克中炎症反应以及血小板活化的重要媒介。因此,我们假设TLR4诱导了肝I / R后血小板ERK5的活化和血小板-CTCs聚集体的形成。我们从肝I/R处理后的血小板特异性TLR4敲除(PF4-TLR4-/-)或TLR4loxp / loxp小鼠中分离血小板。TLR4的基因敲除几乎完全阻断了pERK5 / P70S6K / RAC1的上调(图6A)。为进一步证实TLR4在ERK5信号通路上的作用,我们使用了抗CD41抗体去除上述小鼠的血小板。然后将从野生型和TLR4-/-小鼠获得血小板,移植到血小板缺失的小鼠中,随后进行肿瘤细胞注射和肝I / R处理。与TLR4-/-血小板相比,野生型血小板移植导致磷酸化ERK5以及下游P70S6K和RAC1的表达显著增加(图6B)。这些数据表明血小板TLR4对于手术诱导的ERK5 / P70S6K / RAC1途径的激活至关重要。同时在经肝I / R处理的PF4-TLR4-/-小鼠中GPIIb / IIIa的活化和血小板-肿瘤细胞的聚集也受到抑制(图6C和6D)。上述数据表明血小板TLR4信号是ERK5 / P70S6K / Rac1的关键上游触发因子,对于活化血小板和形成血小板-肿瘤细胞聚集体至关重要。
我们以上数据表明血小板TLR4在ERK5-整合素途径的上游发挥作用。然后我们探讨了TLR4基因敲除是否也存在影响NETs介导的肿瘤细胞捕获的可能性。钙离子载体A23187常被用于刺激中性粒细胞中的NETs形成。如上所述,将经过肝I / R处理后的PF4-TLR4-/-和TLR4loxp / loxp小鼠来源的血小板与MC38细胞预先共培养,然后灌注在中性粒细胞上。当体外动态粘附系统中不存在TLR4时,血小板对NETs介导的肿瘤细胞捕获的促粘附作用降低(图6E)。上述结果与在静态系统中的结果一致(图6F和6G)。以上结果表明血小板中TLR4的基因敲除阻断了NETs介导肿瘤细胞体外捕获中血小板的促黏附作用。
我们还探究了肝I / R后TLR4对NETs形成的影响。结果显示PF4-TLR4-/-小鼠和TLR4loxp / loxp小鼠中循环MPO-DNA复合物的血清水平以及Cit-H3的肺表达无显著性差异(图6H和6I)。上述数据表明血小板TLR4的选择性敲除并不影响NETs的形成,而是通过影响ERK信号通路导致血小板的活化以及NETs介导的CTCs捕获。
肝I / R后激活血小板TLR4-ERK5信号转导的机制仍不明确。我们曾报道过在肝I / R期间肝细胞释放的内源性分子,例如高迁移族蛋白1(HMGB1)和组蛋白,会引起炎症反应。因此,我们紧接着探究外源性HMGB1或组蛋白是否可以触发血小板TLR4-ERK5信号传导。我们发现重组HMGB1和组蛋白均可在体外野生型血小板中显著增加pERK5 / P70S6K / RAC1的表达,而在TLR4-/-血小板中则没有(图6J)。这些结果表明TLR4与其配体的结合在激活血小板ERK5介导NETs捕获肿瘤细胞中起着重要作用。
为了证实上述体外实验结果,我们将从WT或TLR4-/-小鼠获得的血小板转移至血小板缺失的WT小鼠中,随后进行肿瘤细胞注射和肝I / R处理。与野生型血小板相比,TLR4-/-血小板转移显著减少了小鼠肿瘤肺转移(P <0.01)。
为了进一步验证血小板TLR4在术后远处转移中的作用,我们使用了PF4-TLR4-/-和TLR4loxp / loxp小鼠进行体内实验。与TLR4loxp / loxp对照小鼠相比,PF4-TLR4-/-鼠的肺肿瘤显著减少(P <0.01)(图7A和B)。与TLR4loxp / loxp小鼠相比,21天后PF4-TLR4-/-小鼠肺转移负荷显著降低(图7C,7D和7E)。综上所述,我们的研究数据表明血小板TLR4在阻断由NETs介导的肿瘤细胞远处转移中发挥重要作用。血小板TLR4是一个富有前景的肿瘤治疗靶点(图7F)。
目前越来越多的证据表明,宿主免疫细胞在驱动肿瘤进展和远处转移中至关重要。只有一小部分的循环肿瘤细胞能够在脉管系统中幸存,成功地穿过血管转移性传播。应激因素的诱导和远处定植是肿瘤转移的关键。循环肿瘤细胞的行为在很大程度上受其微环境的影响,尤其是循环系统中宿主免疫细胞的影响。本研究描述了先天免疫反应中的两个初始反应细胞,血小板和中性粒细胞如何与循环肿瘤细胞相互作用,以促进在外科手术应激状态下肿瘤的远处转移。这些研究结果证明宿主的系统性生理发生急剧变化会影响包括肿瘤侵袭转移在内的疾病的各个方面。
在这项研究表明手术应激通过ERK5 /整合素GPIIb / IIIa信号激活血小板,导致血小板与CTC聚集增加。先前的研究报道血小板可非常迅速地围绕肿瘤细胞,并且聚集体在肿瘤细胞进入循环后1分钟内定位在该部位,这对于肿瘤早期转移性小灶的形成和随后的转移是必不可少的。在这种“转移前的小灶”模型中,血小板与肿瘤细胞的相互作用会募集粒细胞,从而支持肿瘤转移性播种。整合素、选择素等黏附分子及其配体已被证实可促进血小板与肿瘤细胞的相互作用,其中整联蛋白GPIIb / IIIa在这方面的作用尤为明显。本研究的证据表明,在局部I / R中,血小板中的ERK5立即被磷酸化,其激活增强后通过活化的整联蛋白GPIIb / IIIa促进CTC与血小板的黏附。在肿瘤中其他的信号传导途径和分子可能也参与促进肿瘤细胞与血小板粘附,但来ERK5可能起到了主要作用,其他途径的作用较小。未来用药物抑制ERK5信号传导可能有助于抑制手术引起的血小板活化和随后的血小板肿瘤细胞聚集。
血小板表面的Toll样受体有助于病原体识别,使血小板成为先天免疫的主要组成之一。TLR4膜受体通过与肿瘤细胞释放的HMGB1相互作用,在小鼠B16F10黑色素瘤和Lewis肺癌细胞的实验性肿瘤转移中起主要作用。这些研究表明,靶向血小板TLR4是治疗癌症转移的有效方法。在荷瘤小鼠中观察到TLR4有促进肿瘤转移的作用,这与我们的发现血小板TLR4在接受肝I / R的小鼠转移中起关键作用的结果是一致的。通过使用血小板特异性基因敲除小鼠,我们证实在手术应激后血小板TLR4导致ERK5磷酸化,从而激活血小板,并通过表面整合素GPIIb / IIIa促进其与CTC的聚集。
TLR4在肿瘤细胞和多种宿主细胞中高表达,包括内皮细胞,成纤维细胞,免疫细胞和通过TLR4依赖性趋化因子募集到肿瘤炎症环境中的多种前体细胞。在这些有核细胞中,TLR4可通过NF-κB激活细胞内信号通路,从而导致炎症细胞因子,促存活和迁徙因子的表达上调。TLR4诱导的炎症作用可促进肿瘤生长,血管生成和转移。尽管血小板缺乏基因组DNA,但它们具备RNA剪接能力,可处理常驻前mRNA来存储执行的和合成的免疫调节分子,包括血小板源性生长因子,IL-1β和CCL5,以应对感染性或无菌性损伤。因此,靶向血小板TLR4的治疗可能具有抑制血小板立即磷酸化的潜力,及其活性导致的血小板CTC-粘附特性。
除了与CTC形成聚集体并保护它们免受剪切应力外,活化的血小板还被证明可以直接诱导NETs的形成,并且这些NETs可以增加TRALI小鼠肺部的内皮通透性。NETs在最初被发现时被认为是宿主应对病原体的先天免疫防御机制之一,后来发现其也可促进炎症反应、组织损伤和肿瘤转移。在这项研究中,我们发现在实验性手术应激下,肺微循环中的NETs形成显著增加,血浆中的NETs成分明显增多,从而增强了它们隔离CTC和诱导其远处转移的能力。但是,IR手术前的血小板耗竭并不会影响NETs的形成及其在肺组织中的沉积,这表明活化血小板的促转移作用不会影响NETs的形成。在既往研究中,我们已经阐明了肝脏I / R触发中性粒细胞激活和NETs形成的机制。在肝外科手术期间,从应激的肝细胞(包括HMGB1和组蛋白)中释放与损伤相关的分子模式(DAMP),可通过TLR依赖性途径触发NETs的形成。
NETs是术后脓毒症中肿瘤进展的促进分子。在严重感染的情况下,NETs可以捕获CTC,从而促进其血管内存活和肿瘤细胞早期粘附于远处器官。根据这一思路,NETs在肿瘤转移中的关键作用已在实验性荷瘤小鼠和临床研究中得到证实,在转移性肿瘤的治疗中表现出具有强大潜力。这一现象目前已有的解释机制主要是NETs和CTC之间的粘附是由整合素介导的蛋白质相互作用或NETs DNA与癌细胞上的跨膜蛋白CCDC25的相互作用。在本研究中,我们展示了一种不同的机制:以血小板充当桥梁,在手术应激引起的全身性炎症反应中将CTC连接在肺中沉积的NETs中。
这项研究表明手术应激可通过ERK5 /整合素GPIIb / IIIa信号激活血小板,最终导致血小板-CTC聚集增加。尽管过去已有多份研究表明许多黏附分子包括整联蛋白,选择蛋白及其配体,有助于血小板与肿瘤细胞的相互作用,但这项研究率先证明了血小板活化通路TLR4/ERK5/整合素 GPPIIb/IIIa在促进肿瘤远处转移中发挥了重要作用:肝局部I/R过程中血小板ERK5被迅速磷酸化、并通过活化的整联蛋白GPIIb / IIIa途径赋予CTC黏附性质从而促进肿瘤的远处转移。因此,阻断血小板TLR4或ERK5依赖的血小板-肿瘤聚集体将有望预防由手术引起的远处转移。随着实体瘤的手术切除变得越来越常规,这项可能会为改善切除后患者的临床结局开辟新途径。
不感兴趣
看过了
取消

人点赞

人收藏

打赏
不感兴趣
看过了
取消
©2012-2023 北京华媒康讯信息技术股份有限公司 All Rights Reserved. 注册地址:北京 联系电话:010-82736610
广播电视节目制作经营许可证 —(京)字第 17437号 京海食药监械经营备20200522号
京ICP备12011723号 京ICP证150092号
 京公网安备 11010802020745号
工商备案公示信息
互联网药品信息服务资格证书((京)-非经营性-2020-0015)
京公网安备 11010802020745号
工商备案公示信息
互联网药品信息服务资格证书((京)-非经营性-2020-0015)






您已认证成功,可享专属会员优惠,买1年送3个月!
开通会员,资料、课程、直播、报告等海量内容免费看!

打赏金额
认可我就打赏我~
1元 5元 10元 20元 50元 其它
打赏作者
认可我就打赏我~
扫描二维码
立即打赏给Ta吧!
温馨提示:仅支持微信支付!
已收到您的咨询诉求 我们会尽快联系您




































 010-82736610
010-82736610
 股票代码: 872612
股票代码: 872612




