
正如本文所讨论的,包括粘膜免疫系统、肠道屏障以及肠道微生物群在内的不同胃肠道(GIT)因子,不仅影响GIT的局部疾病,同时也会对全身不同的系统疾病造成影响。
编译:微科盟艾奥里亚,编辑:微科盟茗溪、江舜尧。
微科盟原创微文,欢迎转发转载,转载须注明来源《微生态》公众号。
导读
胃肠道(GIT)不仅会影响GIT的局部疾病,还会对各种全身性疾病造成影响。可同时影响GIT和人体健康和疾病的因素包括1)由肠道相关淋巴组织和固有层组成的粘膜免疫系统;2)由粘液和肠上皮组成的肠道屏障以及3)肠道菌群。包括抗原、免疫调节剂、肠屏障增强剂和肠道微生物群调节剂在内的选择性递送药物,在口服疫苗、免疫耐受、炎症性肠病和其他包括癌症在内的全身性疾病的治疗中表现出可观的结果。然而,GIT的理化和生物学障碍对这些选择性递送药物是否能够成功转运到胃肠道内具有重要影响。随着新型纳米材料的进步,口服纳米医学,由于其不仅克服了这些障碍,而且能够选择性地将药物递送到GIT靶点,使其成为一种具有吸引力的选择方式。在本综述中,我们对GIT因素以及GIT中的理化和生物屏障进行了讨论。此外,我们还介绍了口服纳米医学用于口服疫苗、免疫耐受和抗炎治疗的最新进展。我们还讨论了旨在加强肠道屏障功能和调节肠道菌群和微生物代谢产物的口服纳米医学的最新进展。最后,我们对口服纳米免疫治疗的未来方向提出了看法。
图文摘要
论文ID
原名:Oral nanomedicine for modulating immunity, intestinal barrier functions, and gut microbiome
译名:用于调节免疫、调节肠道屏障功能以及肠道微生物群的口服纳米药物
期刊:Advanced Drug Delivery Reviews
IF:15.470
发表时间:2021.10.26
通讯作者:Yonghyun Lee,James J. Moon
通讯作者单位:韩国梨花女子大学(Ewha Womans University),美国密歇根大学(The University of Michigan)
DOI号:10.1016/j.addr.2021.114021
综述框架图
主要内容
1 引言
胃肠道(GIT)作为人体体内与外部环境直接接触的最大窗口,胃肠道不仅影响胃肠道内的局部疾病(如炎症性肠病、IBD),还会对各种全身性疾病(如糖尿病和癌症)造成影响。因此,针对GIT的治疗方法具有广泛的应用。在开发GIT治疗药物时,应考虑以下因素:1)GIT中由GALT(包括派尔集合淋巴结[Peyer’s patches]和分离的淋巴滤泡在内的肠道相关淋巴组织)、固有层和肠上皮组成的粘膜免疫系统;2)由粘液和肠上皮组成肠屏障以及3)GIT中定植的大量(数量至少在1014以上)且丰富(约10000余种)的肠道共生微生物。由派尔集合淋巴结和淋巴滤泡组成的GALT是诱导针对从肠腔转运的抗原产生免疫应答的主要场所。通过GALT中的诱导过程激活适应性免疫系统后,活化的B细胞和T细胞到达肠道固有层和其他全身粘膜区域,以建立保护性免疫。另一方面,当在固有层中处理抗原时,一般免疫学所产生的结果是对该特异性抗原的免疫耐受。在某些疾病(如IBD、糖尿病和结直肠癌)的状态下,人体的肠屏障功能被破坏,导致对肠道相关抗原的过度免疫反应和粘膜免疫应答失调。值得注意的是,肠道菌群与肠道屏障和粘膜免疫系统紧密相互作用,并提供微生物抗原和代谢产物。此外,肠道菌群的组成和微生态的失调,以及肠道屏障和免疫系统的功能障碍,对各种GIT和全身性疾病的发展、严重程度和治疗都具有很大的影响。因此,开发能够调节GIT因子的GIT靶向治疗药物具有十分重要的意义。
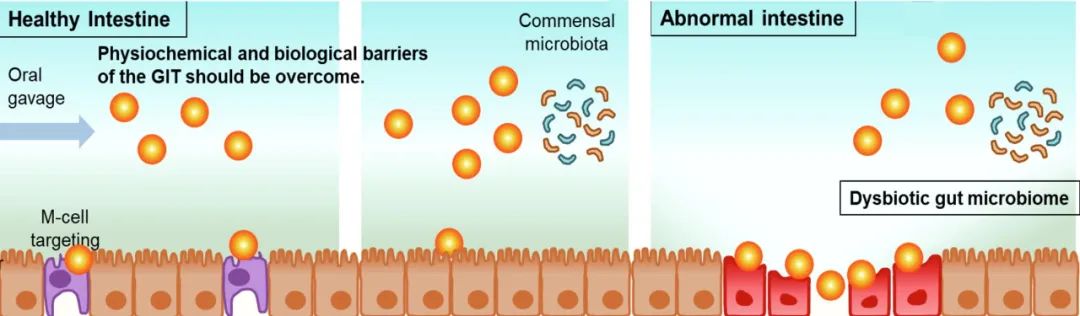
与其他给药途径相比,口服免疫治疗在调节GIT因子方面具有明显优势。口服免疫治疗的主要目标和挑战包括以下几点(图1)。首先,正是由于像派尔集合淋巴结在内的M细胞能够专有的将抗原从管腔转运至淋巴组织的上皮细胞,因此口服疫苗应用中使用的抗原应靶向M细胞。当M细胞将抗原转运自上皮细胞后,派尔集合淋巴结内的免疫细胞将引起抗原特异性免疫应答,并在GI粘膜表面以及全身范围中建立保护性免疫。其次,众所周知固有层摄取的抗原可诱导全身和粘膜免疫耐受,这构成了针对自身免疫性疾病和过敏的实验性口服免疫治疗的基础。再者,固有层的失调与IBD和其他GIT疾病相关;因此,固有层是口服免疫治疗的潜在靶点。第四,肠屏障功能的破坏会导致诸如IBD在内的许多病理发生,因此,恢复和加强肠屏障功能是治疗GIT疾病的可行方法。最后,最新的研究表明,肠道菌群微生态的失调与包括结肠炎、糖尿病和结直肠癌在内的许多疾病有关。因此,恢复健康肠道微生物群不失为是一种能够引起广泛关注的口服免疫治疗的新形式。
图1 口服纳米免疫治疗的生物医学应用。口服纳米药物的免疫治疗包括以下几方面:1)口服疫苗、2)免疫耐受、3)炎症性肠病的治疗、4)增强肠道屏障、以及5)调节肠道菌群。
不同的治疗药物正逐步开发,以满足上述目标。对于小分子药物而言,通过口服给药后,只有一部分药物能够到达GIT的靶点,而大部分通常被机体排泄或被机体吸收,这通常会导致全身的毒性暴露和严重副作用。例如,甲泼尼龙可引起胸腺退化、失眠、抑郁、骨密度降低和满月脸等副作用,类似的例子还有很多,不在一一列举。相比之下,大分子口服给药由于药物的体积较大,全身吸收有限。然而,由于恶劣的GIT条件下(包括pH变化和蛋白水解酶),这些大分子药物容易发生结构变化、降解或是丧失其特有的生物活性。因此,亟需新的口服给药系统,以满足在有效地保护其搭载的药物免受GIT的恶劣条件的同时,选择性地将药物递送到所需的靶位点。
纳米医学由于纳米粒子(NPs)能够保护其搭载的药物分子免受外界胁迫,同时能将药物递送到其靶点,并维持药物释放而备受关注。为了实现这些目标,口服纳米药物应克服GIT中的多种理化和生物学障碍,这些障碍包括胃中的高酸性环境、极端的pH值变化以及沿GIT的蛋白水解酶。对于具有弱酸或碱基团的纳米粒子,应考虑到pH值的变化会影响纳米粒子的可电离基团及其形态的变化。对于靶向派尔集合淋巴结或固有层的NPs,如何克服肠上皮及其粘液分泌层是纳米医学需要考虑的关键障碍。基于以上背景,可以设计能够粘附在粘液层并穿透肠上皮的NPs。另一方面,对于靶向小肠(能够吸收药物的主要场所)的NPs,由于NPs在小肠中的停留时间较短(3-4 h),这使得对于这部分NPs的设计提出了额外的挑战。最后,口服纳米医学的设计标准应考虑健康与病理状态下靶组织的特性。例如,由于IBD的关键特征是上皮细胞炎症的渗漏以及粘液层的损失,用于治疗IBD的口服纳米药物可能被动到达破坏的肠道屏障和固有层。相反,由于大多数共生微生物栖息在肠腔的粘膜层中,对于旨在调节肠道微生物组的口服纳米医学的设计时,应主要考虑靶向粘液层。
本综述论文中,我们首先提出了关键的GIT因素,这些因素包括免疫系统、肠道屏障和肠道微生物群,并回顾了口服纳米药物需要克服的GIT所需要克服的物理化学和生物学障碍。此外,我们还讨论了设计用于口服疫苗、口服耐受和抗炎应用的口服纳米医学的最新进展。最后,我们强调了口服纳米医学的最新进展,其可用于调节肠道屏障功能和肠道菌群,作为治疗各种疾病的新方法。
2 免疫系统、肠道屏障和肠道微生物群之间的相互作用 2.1 胃肠道(GIT)免疫系统
正是由于人体胃肠道(GIT)内含有高达70%的淋巴细胞,使得GIT成为人体最大的免疫器官。肠道免疫系统可分为诱导位点和效应位点。诱导位点包括GALT(如派尔集合淋巴结和孤立的淋巴滤泡)和肠系膜淋巴结(mLN)。主要效应位点是上皮细胞和固有层,这其中包含着大量活化的T细胞和分泌抗体的浆细胞以及包括巨噬细胞和树突状细胞(DC)在内的先天性免疫细胞。胃肠道免疫系统不断受到来自肠腔抗原的攻击,因此必须区分清楚应该对哪些抗原耐受(例如自身抗原、食物、共生微生物),对那些抗原不耐受(例如病原体、毒素)。由于肠道内所含有的高水平的抗炎因子,如IL-10和转化生长因子(TGF)-b等,肠道免疫环境(肠上皮和固有层)通常具有免疫抑制性。
在肠道先天性免疫系统中,肠上皮细胞作为肠道内最外层的哨兵,排列在肠腔内,并作为肠腔内容物和宿主免疫系统之间的物理屏障。肠上皮细胞通过模式识别受体(PRRs)(如Toll样受体[TLRs])从而主动促进先天性免疫应答以及感知微生物。巨噬细胞还具有重要的看家作用,这些作用包括清除凋亡或衰老细胞、组织重塑和维持肠道免疫调节环境。在稳态条件下,Ly6Chigh单核细胞组成性进入肠粘膜(尤其是固有层),并局部分化为抗炎成熟CX3C趋化因子受体1(CX3CR1)high F4/80+巨噬细胞,随后表达清道夫受体和主要组织相容性复合物-II。这些物质对于促炎性刺激响应低下,但对入侵的共生菌或病原体具有高度的吞噬性。此外,固有巨噬细胞通过肠上皮屏障中细胞之间延伸的树突直接采集肠腔内容物,进而协助CD103+ DCs诱导口服耐受。CX3CR1high巨噬细胞同样可以产生大量的IL-10,增强调节性T细胞(Tregs)在粘膜中的二次表达。相应的,Tregs产生的TGF-β可能调节新外渗的单核细胞。固有巨噬细胞还可以通过分泌前列腺素E2(有助于生理性组织重塑)来维持上皮完整性。
炎症或感染可扰乱肠道稳态。上皮细胞在PRRs感染的初始定植和感知中发挥重要作用。当炎症或感染扰乱体内平衡时,单核细胞分化的正常模式被破坏,这会导致促炎效应因子Ly6Chigh单核细胞和CX3CR1int巨噬细胞的蓄积。巨噬细胞通过TLRs感知扰动,从而激活细胞内促炎信号。固有单核细胞通过产生巨噬细胞源性的趋化因子来促进中性粒细胞的招募。血液循环中的中性粒细胞可感知化学引诱物的梯度,并从血管内皮渗出到达肠固有层。中性粒细胞具有一定的防御机制,这些机制包括如烟酰胺腺嘌呤二核苷酸磷酸、氧化酶介导的活性氧簇以及抗菌肽等。中性粒细胞还可促进其他免疫细胞的募集,并促进粘膜愈合。在炎症过程中,固有的CX3CR1high巨噬细胞保留其抗炎特性,如分泌IL-10。经典的单核细胞和固有CX3CR1high巨噬细胞通过调控和清除活化的有害中性粒细胞从而发挥调节作用。
扩大先天响应会对宿主产生不良的影响。病理状态下肠道中促炎性单核细胞、活性中性粒细胞和其他先天性免疫细胞的过度募集和蓄积与粘膜损伤和衰弱性疾病症状有关。此外,活性先天性免疫细胞产生的肿瘤坏死因子(TNF)-α在保护宿主方面发挥着重要的作用,但当TNF-α大量产生或长期产生时,会导致包括DNA损伤在内的组织损伤。由于大多数细胞表达TNF-α受体,这会导致活性氧积累的增加、趋化因子水平的升高及其他促炎反应。TNF-α具有混杂效应,使其成为IBD患者的有效治疗靶点。
作为适应性免疫应答的主要诱导位点,GALT和mLN携带记忆B细胞和T细胞,随后能够通过淋巴系统迁移到粘膜效应位点(肠道粘膜和全身粘膜效应位点),进而发挥粘膜免疫应答响应(图2)。在诸如派尔集合淋巴结和分离的淋巴滤泡在内的GALT组织中,覆盖在派尔集合淋巴结和淋巴滤泡上层的特异性M细胞在抗原的跨细胞转运中发挥着关键的作用。M细胞能够将抗原传递给树突状细胞(DC细胞)。GALT中的DC也可通过其延伸的树突直接从管腔中摄取抗原,随后,DC处理抗原并在其表面传递抗原片段,激活初始CD4+ T细胞。进一步,CD4+辅助性T细胞与抗原特异性B细胞相互作用,导致B细胞转变为免疫球蛋白分泌细胞。活化的B细胞离开派尔集合淋巴结和淋巴滤泡,到达远端效应位点(即全身粘膜和肠道效应位点),导致其分化并成熟为浆细胞。另外,携带抗原的DC本身也可以直接迁移至淋巴结,与生长中心相互作用,并激活体液和细胞应答,导致活化的免疫细胞(B细胞和T细胞)迁移至远端效应位点。但目前仍不清楚M细胞介导的抗原摄入到GALT中这一过程是否在诱导对可溶性抗原的口服耐受中具有重要作用。此外,虽然由派尔集合淋巴结和独立的淋巴滤泡介导的抗原摄取在口腔耐受中只起到轻微的作用,但它们可能在调节肠道微生物群的免疫应答中更为重要。在某些情况下,低剂量抗原的多次给药或无促炎危险信号的高剂量抗原可能引起GALT的口服耐受,关于这一点,在开发口服疫苗和口服耐受免疫治疗时应值得考虑。
图2 胃肠道中的免疫系统。(I)代表覆盖在GALT上皮细胞中的特异性M细胞介导抗原跨细胞转运到树突细胞中;(II)代表B细胞被激活为免疫球蛋白分泌细胞,随后迁移到固有层和全身黏膜效应部位,并分化为IgA分泌浆细胞(Ⅲ);(IV)代表携带抗原的树突细胞迁移到mLN并激活B细胞和T细胞,随后通过淋巴系统迁移到远端的效应位点(V);(VI)代表抗原可通过上皮紧密连接扩散、跨细胞途径的跨上皮细胞转移、外泌体介导的递送或从CX3CR1high巨噬细胞捕获等方式被树突细胞识别;(VII)代表携带抗原的CD103+树突细胞激活了口服耐受,并诱导Tregs,随后导致其播散到远端效应位点(VIII);(IX)代表抗原也可到达肝脏和外周淋巴结;(X)代表口服免疫疗法可触发保护性免疫或耐受性免疫,但这取决于靶细胞和局部信号。
固有层(肠道效应位点)含有抗原特异性粘膜效应细胞,这些细胞包括产生免疫球蛋白(Ig)A的浆细胞以及记忆B细胞和T细胞,并具有维持体内平衡的粘膜保护活性。例如,来自浆细胞的IgA通过聚合Ig受体跨上皮转运。分泌型IgA抗体可防止病原体在粘膜表面附着和定植。此外,有助于宿主防御固有层病原体定植的其他重要效应机制包括局部产生的IgM和IgG以及粘膜细胞毒性T淋巴细胞。在IBD炎症条件下,固有层会发生改变,其主要表现为免疫细胞活化频率的增加、肠屏障的渗漏、病原菌或肠道菌群的渗透以及免疫应答失调。正如在IBD患者中所观察到的那样,上述固有层的改变会使得固有层成为容易发生组织损伤的发炎环境。
另一方面,固有层也可作为口服耐受的诱导部位。固有层中CD103+ DCs摄取抗原可诱导口服对可溶性抗原的耐受。DCs可通过以下途径来对抗原进行识别:1)通过上皮紧密连接扩散;2)通过跨细胞途径的跨上皮细胞转移;3)外泌体介导的递送;以及4)从CX3CR1high巨噬细胞中捕获。携带抗原的CD103+ DC从固有层迁移至mLNs,激活口服耐受。在mLN中,从维生素A中产生的维甲酸(RA)能够诱导抗原特异性T细胞上的肠道归巢受体α4β7整合素和C-C基序趋化因子受体9的表达,以及Foxp3+ Tregs的TGF-β依赖性分化。Tregs重新进入肠固有层,并在CX3CR1high巨噬细胞产生的IL-10的影响下发生继发性扩增。随后,Tregs通过淋巴循环或体循环进入其他免疫区域,建立全身口服耐受。摄取到派尔集合淋巴结或固有层的抗原也可能到达肝脏,肝脏中的肝窦内皮细胞、耐受的常规DCs或浆细胞样DCs可进一步诱导全身耐受。当抗原到达外周淋巴结,同时在没有共刺激的情况下由固有DC呈递时,也可能发生全身耐受。
2.2 肠道屏障
肠道屏障由单层肠上皮细胞组成,参与诱导和维持先天性免疫。除此之外,肠道屏障还包括杯状细胞、潘氏细胞、M细胞、肠上皮干细胞和肠内分泌细胞在内的其他细胞。其中,肠细胞、杯状细胞和M细胞在肠道保护、转运和免疫中发挥关键作用。GIT上皮上的黏液层能够保护机体免受致病威胁的侵袭(图3)。粘液主要由杯状细胞分泌的粘蛋白组成,其本质上是由蛋白质、碳水化合物、脂质、盐以及抗体等混合组成一种水凝胶(95%的水)。粘液的屏蔽和润滑功能在维持肠道稳态中起着至关重要的作用。粘液层直接与肠道菌群连接,并为微生物的多糖结合组分提供附着位点,从而影响微生物的定植。粘蛋白多糖也可作为粘液相关细菌(即所谓的“粘液溶解细菌”)的营养物,促进细菌的繁殖。基于上述的这些作用,塑造了组成我们肠道菌群的各种共生微生物。此外,诸如乳杆菌属(Lactobacillus spp)、长双歧杆菌(Bifidobacterium longum)、罗伊氏乳杆菌(Lactobacillus reuteri)以及嗜黏蛋白阿克曼菌(Akkermansia muciniphila)在内的一些共生微生物,可促进粘蛋白的产生并增加粘液层厚度,进而提供额外的肠道屏障,阻止了病原微生物的粘附,从而为机体对抗病原微生物提供保护作用(图3)。粘液层的粘度也限制了微生物的运动,从而保护粘液层下的上皮细胞免受微生物的侵害。总体而言,粘液屏障的完整性对于GIT的保护功能至关重要。
图3 维持肠屏障功能。肠屏障由肠上皮细胞和粘膜层组成。肠上皮细胞通过一系列细胞间紧密连接连接连接,负责肠屏障功能。粘膜和肠道菌群层保护机体免受病原体的侵害。肠屏障破坏与包括IBD、高血糖、感染以及癌症在内的各种疾病有关。
肠道屏障功能对于维持组织稳态至关重要。单个上皮细胞之间的连接由一系列细胞间紧密连接组成,这些链接方式包括连接粘附分子、紧密连接蛋白、claudin以及tricelluin。紧密连接是沿侧面的顶端连接,直接负责肠道屏障功能。当细胞与细胞间连接的完整性被破坏时,病原菌和分子可能会无限制地穿过上皮层。此外,粘液层和肠道菌群对肠屏障功能也十分重要。肠上皮屏障不断受到肠道菌群、食物和食物相关微生物的挑战,肠屏障功能、上皮完整性和细胞-细胞连接的失调与多种病理状态、以及诸如炎症性肠病(IBD)、自身免疫性疾病和全身感染在内的疾病息息相关(图3)。IBD易感基因C1orf106(1号染色体开放阅读框106)的失活可降低肠道屏障功能,从而触发肠道炎症和IBD。C1orf106蛋白在维持成熟上皮适当的cytohesin 1蛋白浓度中具有重要作用。Cytohesins作为Ras鸟苷三磷酸酶ARF(ADP核糖基化因子6)的激活剂,其指导了钙粘蛋白的内吞内化。ARF6活性的下调在维持紧密连接的稳定性中具有重要意义。具体而言,C1orf106的耗竭导致细胞素的异常积累以及ARF6的过度激活,进而增加钙粘蛋白内吞和紧密连接的通透性。此外,细菌成分、细菌碎片和其他抗原分子可以通过渗漏的紧密连接结构引起免疫反应、炎症和组织损伤。例如,致病菌鹑鸡肠球菌(Enterococcus gallinarum)可诱导肠屏障缺陷并转移至淋巴结以及肝脏,引发自身免疫性疾病--系统性红斑狼疮。此外,与糖尿病和其他代谢综合征相关的高血糖症可破坏肠道屏障,导致肠道炎症和感染。慢性高血糖通过肠上皮细胞中葡萄糖转运蛋白GLUT2的代谢和转录重编程影响屏障功能,导致细菌副产物播散和全身炎症。总体而言,这些研究表明肠屏障功能在预防IBD和其他全身性疾病中的关键作用。然而,肠屏障渗漏的机制尚不清楚,需要进一步研究。
2.3 肠道微生物
人体体内栖息着数以万亿的微生物,这些微生物包括细菌、真菌以及病毒。据估计,约有超过1014个细菌菌种定植于胃肠道内(GIT)。肠道菌群通过在粘液层中形成额外的屏障来补充肠道屏障,将病原体、颗粒和污染物与内部环境分开。值得注意的是,肠道微生物群与我们的免疫系统和肠道屏障建立了共生关系。作为宿主-微生物组共生的一个例子,一些诸如嗜黏蛋白阿克曼菌(Akkermansia muciniphila)的微生物组以粘蛋白层为食,而共生微生物是粘液层岩藻糖基化及其完整的肠屏障功能所必需的。肠道菌群还参与先天性免疫系统的成熟和功能,这些功能包括抗菌肽的分泌,以及IL-22、IL-17和IL-10的产生。因此,肠道微生物-宿主相互作用在免疫系统的发育、成熟和维持中起着至关重要的作用(图4)。
图4 肠道微生物群在各种疾病中的作用。益生菌肠道菌群与包括感染、炎症相关疾病和癌症在内的各种局部和全身疾病有关。
肠道菌群多样性是指个体中存在的不同共生微生物种类的数量。肠道微生态失调的典型特征是微生物多样性降低和常驻微生物种类的实质性转变。在营养不良的情况下,肠道菌群的失调增加了肠道的易感染性,并导致疫苗应答受损。微生态失调的典型例子是艰难梭菌(Clostridioides difficile)感染,其可快速破坏肠道微生物群落,导致初级胆汁酸和简单碳化合物的富集,最终导致艰难梭菌的繁殖。其他病理状态,如多发性硬化(MS),也与肠道菌群共生相关。MS的特征是免疫介导的中枢神经系统髓鞘的破坏,已有的研究表明,遗传易感染的个体的肠道菌群会影响这一过程。抗生素治疗耗尽肠道共生细菌可改善小鼠实验性自身免疫性脑脊髓炎(EAE)的发生。相比之下,腹膜内抗生素治疗对肠道菌群的影响以对EAE的诱导作用很小,这表明EAE的诱导受肠道微生物组的影响。此外,MS患者的流行病学研究表明,MS患者肠道特定的细菌分类群发生了变化。综上所述,这些研究表明肠道菌群的失调与各种GIT疾病之间的相互关联;因此,使用抗生素、粪便菌群移植(FMT)或补充益生菌等方式可作为艰难梭菌感染、IBD和MS的预防措施或潜在治疗措施。
肠道微生物群在癌变中的作用也逐渐受到研究学者的重视。目前已知的致癌性肠道细菌肠道沙门氏菌(Salmonella enterica),幽门螺杆菌(Helicobacter pylori)等。肠道致癌细菌通过触发局部慢性炎症促进癌变,一些细菌(如幽门螺杆菌)对粘膜细胞具有直接遗传毒性作用。此外,临床前和临床研究表明,肠道微生态的失调是结直肠癌(CRC)的致癌驱动因素。包括脆弱拟杆菌(Bacteroides fragilis)、具核梭杆菌(Fusobacterium nucleatum)、大肠埃希菌(Escherichia coli)和空肠弯曲菌(Campylobacter jejuni)在内的一些细菌与CRC癌变和随后发生的癌细胞转移相关。肠道菌群失调也与肝细胞癌(HCC)和乳腺癌有关。肠道细菌成分、代谢产物和副产物可通过门静脉系统转运至肝脏,引起炎性改变和肝毒性。同样,胆汁酸的微生物衍生物与致癌作用有关。由于未吸收的胆汁酸具有抗菌性,细菌可将胆汁酸转化为具有致癌特性的次级胆汁酸(脱氧胆酸和石胆酸)。肠道菌群的失调也可能通过肠道菌群介导的对雌激素代谢、能量代谢和肥胖的影响从而促进乳腺癌形成。
肿瘤微环境中的细菌定植也可通过与癌细胞的直接相互作用或通过诱导炎症或免疫抑制直接影响癌细胞的生长和免疫应答。有的研究表明,癌症患者不同器官之间的微生物群的具有特异性。值得注意的是,肿瘤内细菌与肠道细菌群之间有很强的相关性。最近在胰腺癌中的研究发现,胃上皮层的完整性受损可能会导致细菌易位到远端肿瘤。有趣的是,肿瘤定植细菌也可能导致化疗耐药。例如,胰腺肿瘤中的支原体(Mycoplasma)通过将吉西他滨代谢为非活性形式从而使得其产生耐药性。这些研究表明了肠道微生物组的促肿瘤特性。
相反,也有研究表明“有益”肠道菌群的抗肿瘤作用。广谱抗生素对肠道菌群的扰动使得患者对免疫检查点阻断治疗中的结局产生负面影响,从而突出了共生菌群在癌症免疫治疗过程中调节免疫应答的重要性。此外,临床前研究表明,共生细菌的给药和随后的肠道定植,增强了免疫检查点阻断治疗的抗肿瘤疗效。此外,环磷酰胺的抗肿瘤作用部分归因于肠道菌群的变化。因此,通过清除‘有害’致癌细菌或移植具有抗肿瘤特性的‘有益’微生物来调节微生物群可能是未来针对于癌症治疗的新策略。
3 用于固有层免疫系统调节的口服纳米药物
由于效应免疫细胞位于固有层,因此目前试图寻求能够调节固有层免疫细胞的方法来治疗IBD和其他GIT相关疾病。例如,固有层中巨噬细胞和中性粒细胞浸润是IBD的标志。因此,固有层中的巨噬细胞和中性粒细胞是IBD的潜在治疗靶点。如果可以通过NPs将药物特异性地递送到靶免疫细胞,这将最大限度地提高药物的疗效,同时最大限度地降低毒性。为了实现这一目标,NPs应该克服GIT中的生物和理化障碍--包括pH变化、沿GIT的蛋白水解酶等。除此之外,药物还需要克服穿透粘膜层和肠上皮层的额外障碍,才能更大的发挥治疗效果。除此之外,还需要考虑到NPs在小肠(3-4h)和结肠(1-2天)中的滞留时间上的差异。值得注意的是,由于IBD中与肠道炎症相关的粘液层的裸露以及受到破坏的肠上皮层,这可能通过渗漏的上皮将载药的NPs被动递送至炎症部位(表1)。
表1 设计GIT靶向口服NPs时需要考虑的关键因素。
对于IBD的治疗,目前已开发出能够靶向激活巨噬细胞表面受体过表达的NP基药物。例如,在炎症条件下,甘露糖受体和巨噬细胞半乳糖型凝集素在活化的巨噬细胞上高表达。目前已经开发出靶向巨噬细胞的NPs(图5)。这些纳米粒子由甘露糖基化生物还原阳离子聚合物、三磷酸钠和TNF-α siRNA组成,表现出高效的巨噬细胞靶向能力,同时未表现出明显的上皮细胞摄取现象。这导致葡聚糖硫酸钠(DSS)诱导的结肠炎小鼠模型具有较强的抗炎活性。口服这些纳米粒子可改善DSS诱导的结肠炎。此外,TNF-α-siRNA负载的聚乳酸-(聚乙二醇)(PEG)NPs与F4/80抗体的Fab’部分共移植,能够靶向巨噬细胞,并且在DSS诱导的结肠炎模型中具有良好的治疗效果。然而,在GIT转运过程中有可能降解接枝配体抗F4/80抗体;因此,将聚乳酸-PEG NPs装载到结肠特异性的生物可降解水凝胶(壳聚糖/藻酸盐)中,从而保护NPs上的Fab’片段,进而改善NPs的菌落特异性递送和治疗效果。
图5 靶向固有层的口腔纳米药物。TPP(三磷酸钠)-PPM(一种甘露糖基化生物还原阳离子聚合物)/siRNA NP(NP)材料的形成以及巨噬细胞靶向传递过程,以及siRNAs释放到细胞质中过程的示意图。
IBD炎症肠道中的Ly6C+炎性白细胞也可作为纳米医学的靶点。携带IL-10 mRNA的脂质纳米粒子通过靶向抗Ly6C抗体,在Ly6C+炎性白细胞中实现了IL-10的选择性表达,在DSS结肠炎模型中产生了可观的治疗效果。其他siRNA负载的NPs已被报道靶向炎性白细胞。上皮细胞和免疫细胞可增加IBD中细胞周期蛋白D1(CyD1)的表达。在DSS结肠炎模型中,携带siRNA CyD1并靶向炎性白细胞的NPs可抑制CyD1 mRNA和炎症反应。同样,抗Ly6C抗体移植的脂质纳米粒子携带针对免疫调节蛋白干扰素调节因子-8的siRNA,靶向炎性Ly6C+白细胞,阻断干扰素调节因子-8 mRNA,显著降低单核吞噬细胞的分化、极化和活化。
探索天然存在的胞外囊泡(如来源于可食用植物的外泌体)是细胞特异性主动靶向方法的另一个新兴领域。例如,磷脂酰乙醇胺和富含磷脂酰胆碱的葡萄柚来源的可食用纳米囊泡通过网格蛋白依赖性途径被肠巨噬细胞摄取,并且由于磷脂酰乙醇胺和磷脂酰胆碱在外层富集而导致微胞饮作用。因此,载甲氨蝶呤(methotrexate)的纳米囊泡表现出巨噬细胞靶向能力,并具有改善DSS诱导的小鼠结肠炎的功效。此外,TGF-β1基因修饰的DC来源的外泌体诱导CD4+ Foxp3+ Tregs,同时减少GIT炎症部位的辅助性T细胞(Th17),从而对DSS诱导的小鼠结肠炎发挥疗效。
除IBD外,GIT靶向NPs(携带抗原的NPs与免疫调节药物,如维生素D和雷帕霉素)已被证明可诱导耐受性免疫应答。这些诱导耐受性的纳米粒子可以阻止抗蛋白药物抗体反应,并可用于治疗包括关节炎、过敏和糖尿病在内的各种疾病。
4 用于调节肠道屏障的口服纳米药物
肠道屏障的破坏与炎症和全身性感染息息相关,肠道屏障的破坏会导致细菌或其副产物渗漏到下层组织和体循环中。因此,恢复破坏的肠屏障完整性将有助于治疗GIT相关疾病。例如,在T细胞介导的急性腹泻小鼠模型中,调控肌动球蛋白收缩力或调控内吞作用的药物减轻了急性腹泻的症状。此外,调节紧密连接蛋白的C1orf106的稳定可能为改善IBD的肠屏障功能提供一种治疗策略。此外,修复上皮层可加强肠屏障功能,这种方式同样可作为IBD防治的新途径。同时,肠细胞的分化强化了肠隐窝,从而为改善肠屏障功能提供了另一个潜在的靶点。
旨在克服GIT生物和理化屏障的口服纳米医学可能为将药物递送至肠上皮和固有层提供新的方法(表1)。作为抗IBD的潜在治疗方法,基于NP的各种策略以被研究,用于调节肠屏障功能。粘膜粘附纳米粒子可以将药物递送到小肠或大肠的粘液层。由于粘蛋白由亲水性组分组成,而纳米粒子含有诸如羧基或羟基在内的亲水性官能团,这可以促进粘蛋白和纳米粒子之间形成氢键。此外,可以通过静电作用,使得NPs靶向到达粘液层。例如利用粘液中的阴离子与NPs表面的阳离子聚合物(如壳聚糖)之间的电荷相互作用。合成聚合物(如丙烯酸衍生物/聚丙烯酸酯)和天然聚合物(如透明质酸、纤维素衍生物、壳聚糖、藻酸盐和果胶)已被证明非特异性粘附粘蛋白,可用于粘液靶向治疗。
鉴于IBD患者炎症区域的粘液层较薄,因此在设计用于治疗IBD的粘液靶向NPs时我们应该考虑到这一点。NP平台的大小和电荷都将影响其对炎症上皮的靶向作用。已有的研究表明,由于小粒径的NPs允许其与IBD受损上皮上表达的带正电荷的蛋白发生相互作用,这使得直径小于200 nm且表面带负电荷的纳米粒子在IBD治疗中表现出更好的组织穿透活性。为了特异性靶向炎症上皮,目前探索了针对炎症上皮上上调的受体/分子的主动靶向策略。例如,肽转运蛋白1(一种寡肽转运蛋白)在IBD患者的结肠上皮中过表达。KPV(Lys-Pro-Val)肽与肽转运蛋白1具有高亲和力,并通过恢复发炎上皮功能发挥抗炎作用。基于KPV的纳米粒子通过加速粘膜愈合从而减轻炎症。与之相似的,IBD、结肠腺瘤和结肠腺癌患者的肠道粘膜组织和结肠微血管中的细胞间粘附分子1(ICAM-1)存在显著上调的现象,这同样可作为口服纳米医学的靶点。目前已经报道了抗ICAM1抗体包被的聚苯乙烯纳米粒子。但在口服给药后,这些NPs主要沉积在胃和十二指肠中,NPs上大约60%的抗icam-1抗体被GIT酶迅速降解,这为成功的口服给药提出了挑战。转铁蛋白受体(TfR)是在炎症结肠肠上皮细胞基底外侧和顶侧膜中过表达的另一个靶点。因此,ICAM-1和TfR靶向策略有可能将药物靶向炎症组织,恢复肠屏障功能,但这仍需要更多的研究来保护靶向配体在GIT中不被降解。
鉴于恢复肠道屏障功能和调节活化免疫细胞在IBD的管理中具有十分重要的意义,因此靶向结肠上皮和活化免疫细胞上过表达的分子可能表现出协同治疗活性。例如,CD98和CD44在结肠上皮过表达,且属于活化免疫细胞,因此可作为潜在的靶标。目前已经开发出一种可用于递送表面涂有单链CD98抗体的CD98 siRNA负载NPs的口服水凝胶(图6),其在小鼠结肠炎模型中,能够靶向CD98过表达的巨噬细胞和炎症上皮,有益地调节其功能。此外,透明质酸也被用作纳米粒子上的主动靶向配体。透明质酸是由N-乙酰葡糖胺和D-葡萄糖醛酸单元组成的天然多糖,其无毒性且可生物降解。携带CD98 siRNA和抗炎性姜黄素的透明质酸功能化聚合物NPs可保护粘膜层,调节活化巨噬细胞的炎症功能,减轻粘膜炎症。同时,在DSS诱导的结肠炎小鼠模型中,透明质酸负载KPV的聚合物NPs加速了粘膜愈合并减轻了小鼠炎症状态。
图6 口服纳米药物可改善肠道屏障功能。a代表单链CD98抗体功能化siRNA负载NPs的自组装过程;b代表负载了NPs的scCD98(绿色)对体外结肠上皮细胞(上图)和巨噬细胞(下图)的特异性;c-d代表基于体重变化(c)、CD98 mRNA浓度(d左)和TNF-α mRNA浓度(d右)探究了siCD98 NPs的体内治疗疗效。
IBD患者的肠黏膜表现为活性氧的过度产生和抗氧化剂的失衡,从而导致黏膜氧化损伤。为了解决这些问题,MeO-PEG-b-PMOT两亲性嵌段共聚物基的胶束在疏水段具有稳定的氮氧化物自由基。这些胶束显著聚集在结肠粘膜区域,尤其是炎症部位,保护上皮区域免受氧化损伤,并在DSS结肠炎小鼠模型中表现出强效的治疗效率。
综上所述,这些研究表明,口服纳米医学可能调节局部GIT疾病的诱导、发展和严重程度,为治疗其他全身性疾病提供一条新途径。
5 用于肠道微生物群调控的口服纳米医学
肠道微生物群与免疫激活和免疫耐受以及各种病理状态都有着错综复杂的联系。因此,目前的研究正在探索能够调节肠道微生物群的策略,尤其是通过肠道靶向NPs和微粒来调节肠道微生物群。肠道微生物群靶向NPs也应该克服GIT的生物和理化障碍。但与上述不同,这些NPs会靶向于驻留在肠腔中的共生微生物,因此不需要穿透肠上皮(表1)。
目前报道了诸多纳米医疗系统,这些系统都能够调节肠道菌群而作为抗癌平台。例如,与人参皂苷Rg3(NpRg3)共轭的Fe@Fe3O4 NPs可被用于HCC的治疗。人参皂苷Rg3被用作自噬抑制剂和阿霉素介导的抗HCC抗癌活性的增敏剂。这种纳米复合材料改变了肠道微生物群的组成,提高了拟杆菌门(Bacteroidetes)和疣微菌门(Verrucomicrobia)的相对丰度,同时降低了厚壁菌门(Firmicutes)的相对丰度。同时,NpRg3显著降低HCC发生过程中的重要代谢产物3-吲哚丙酸和尿素,同时增加游离脂肪酸。因此,NpRg3通过重塑不平衡的肠道菌群及其代谢物抑制了HCC的生长和肺转移。此外,肠道微生物靶向策略已被探索用于CRC的治疗。目前设计出一种可以靶向具核梭杆菌(F. nucleatum,一种致癌细菌)和丁酸梭菌(C.butyricum,一种益生菌)的噬菌体引导的伊立替康-葡聚糖杂交的纳米系统以治疗CRC。在该系统中,利用噬菌体特异性裂解具核梭菌(该细菌可诱导化疗耐药和免疫抑制)。此外,在纳米系统中引入右旋糖酐促进丁酸梭菌增殖,抑制结直肠癌生长并通过产生短链脂肪酸诱导抗肿瘤免疫应答。这种联合策略对CRC表现出协同抗癌活性。除此之外,丁酸梭菌(用作益生菌)和双氯芬酸(用作化疗药物)共同负载的右旋糖酐胶囊能够协同发挥抗肿瘤活性从而治疗CRC(图7)。作为一种替代策略,具有内在抗菌特性的纳米粒子已被用于减少有害细菌,被视为是一种新形的抗癌治疗。已有的研究表明,具有抗菌特性的银纳米粒子可减少与癌症治疗耐药相关的肿瘤内细菌,并在小鼠中对胰腺癌发挥抗肿瘤作用。另一项研究发现,一例头颈部鳞状细胞癌患者在每天使用自制的银纳米粒子3个月后,其癌症的放射影像持续缓解。基于这一示例以及已发表的临床前研究,作者建议应进一步探索银纳米粒子对头颈癌的安全性和有效性。
图7 口服纳米医学用于调节肠道微生物群以用于抗癌治疗。a代表葡聚糖包裹的益生菌(C. butyricum)来调节肠道菌群,抑制结肠癌;b代表微生物代谢产物之一的短链脂肪酸调节肠道菌群,抑制肿瘤生长;c-e代表载双氯芬酸孢子-dex(DC@spores-dex)在小鼠皮下(c,d)或原位CT26肿瘤(c,e)的治疗效果。
纳米医学以外的平台技术也可以调节肠道菌群,作为一种潜在的抗癌疗法。我们最近筛选了FDA公认安全的成分清单,发现口服菊粉(菊苣根和菊芋中发现的一种多糖膳食纤维)可提高免疫检查点阻断剂治疗的抗肿瘤疗效。基于此,我们设计了“结肠保留”菊粉凝胶,以靶向结肠中普遍存在的“有益”共生微生物。在荷瘤小鼠中口服菊粉凝胶治疗增加了在T细胞免疫中的发挥“有益”作用的关键共生微生物(例如Akkermansia、乳酸杆菌[Lactobacillus]、罗氏菌属[Roseburia])的及其代谢物(短链脂肪酸)的相对丰度。这导致IFN-γ+CD8+ T细胞的记忆回忆反应增强,并在肿瘤微环境中建立类干细胞的Tcf1+PD-1+CD8+ T细胞。在多种鼠肿瘤模型中,口服菊粉凝胶与全身免疫检查点阻断剂治疗实现了协同作用,从而突出了靶向肠道微生物群改善癌症免疫治疗的潜在益处。
肠道炎症通常与疣微菌门(Verrucomicrobia)、拟杆菌门(Bacteroidetes)和厚壁菌门(Firmicutes)群落的显著减少相关,尤其以嗜黏蛋白阿克曼菌(Akkermansia muciniphila)、Clostridium XIV α、乳酸杆菌(Lactobacillus)、Clostridium coccoides和肠道柔嫩梭菌(Clostridium leptum)等细菌菌种丰度的降低相关。肠道炎症还与放线菌门(Actinobacteria)和变形菌(Proteobacteria)群落的显著增加相关,尤其是肠杆菌科(Enterobacteriaceae)。因此,各种益生菌策略已被用于探索作为抗IBD的潜在疗法。例如,口服可表达抗炎性白细胞介素-10(IL-10)的益生菌菌株乳酸乳球菌(Lactococcus lactis)可恢复肠道稳态功能,并防止小鼠在IL-10-/-小鼠中发生DSS诱导的结肠炎。此外,其他工程菌株如干酪乳杆菌(Lactobacillus casei)、植物乳球菌(Lactococcus plantarum)和Streptococcus gordonii等其他益生菌也被研究可用于治疗IBD。这些工程益生菌被认为定植于肠道,产生具有有益作用的重组蛋白,从而影响微生物生态系统,进而重塑微生物群结构。
基于可食用益生元的植物源纳米粒子可用于靶向微生物。研究发现,口服姜黄脂质NPs(GDLPs)能够以脂质依赖性方式靶向鼠李糖乳杆菌GG(Lactobacillus rhamnosus GG,LGG)。携带mdo-miR7267-3p microRNA的GDLPs介导靶向LGG单加氧酶,增加吲哚-3-甲醛,诱导IL-22的产生,导致DSS结肠炎小鼠模型肠道屏障功能改善。这些发现表明,可食用的基于植物的NPs可以用来靶向微生物并缓解IBD的炎症。
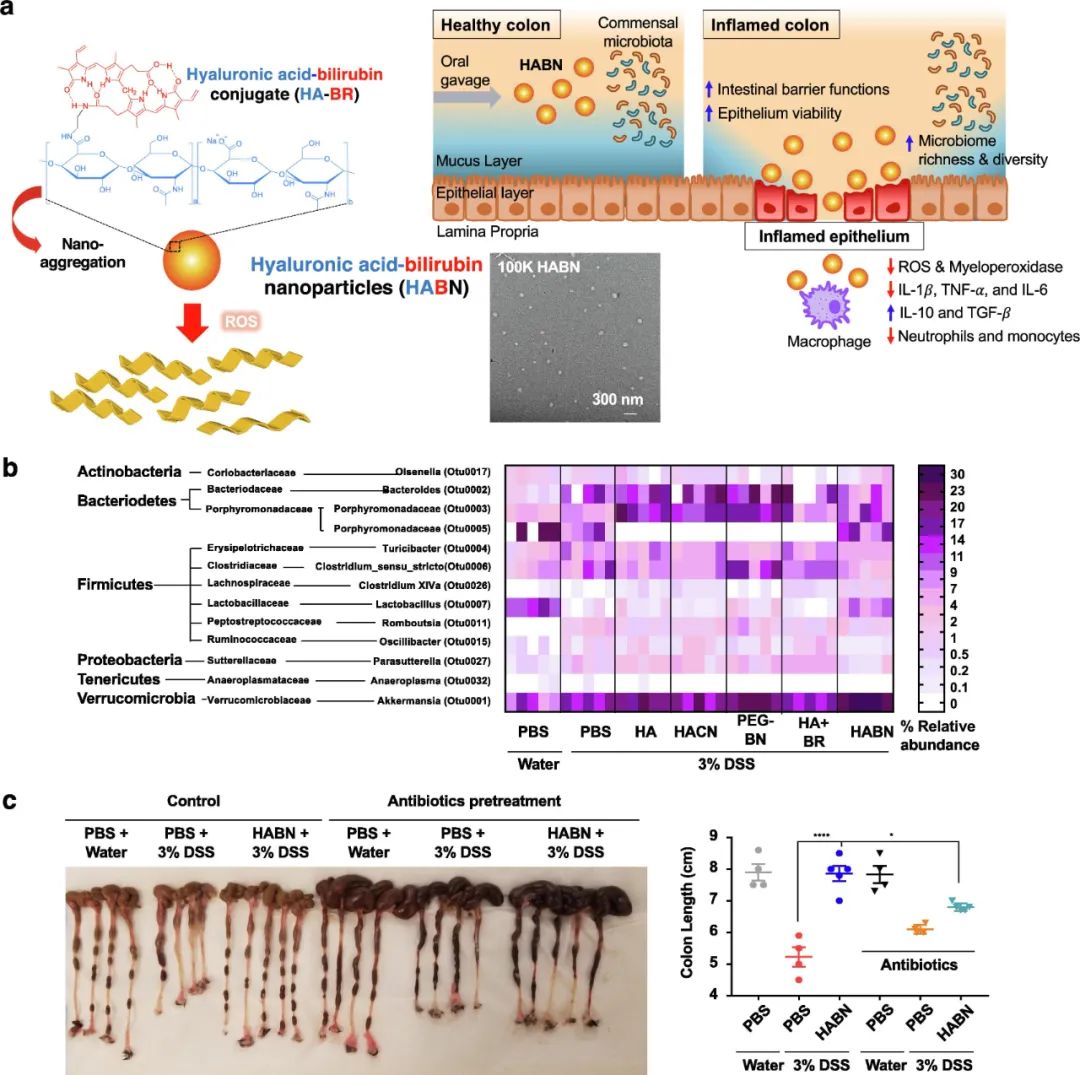
IBD的发病机制与肠道屏障功能破坏、肠道微生物菌群失调以及随后对肠道共生细菌的粘膜免疫应答失调相关。因此,同时针对所有这些因素可能会改善IBD管理的结果。最近,我们研究发现,透明质酸-胆红素NPs(HABN)可以调节肠道微生物群,恢复肠道屏障功能,同时在DSS结肠炎的小鼠模型中发挥抗炎作用(图8)。HABN经口给药能够改变肠道微生物群,增加了嗜黏蛋白阿克曼菌(已知可诱导保护性肠屏障)、Clostridium XIV α(已知可诱导调节性CD4 T细胞)和乳酸杆菌(已知具有抗炎作用)的多样性和相对丰度(图8b)。值得注意的是,抗生素介导的肠道微生物耗竭消除了HABN的抗炎作用,这一结果表明肠道微生物群在基于HABN的治疗中的关键作用(图8c)。除此之外,HABN可恢复肠上皮的保护性肠屏障功能和抗炎免疫应答,从而改善DSS诱导的结肠炎(图8c)。
图8 用于通过改变肠道微生物群从而治疗IBD的口服纳米医学。a代表透明质酸和胆红素结合物(HA-BR)自组装形成的透明质酸-胆红素NPs(HABN)的示意图及其TEM图像。HABN在发炎的结肠中积累,通过靶向调节免疫系统、肠道屏障和肠道菌群发挥抗急性结肠炎的治疗作用。b代表口服HABN可调节DSS结肠炎小鼠的肠道微生物群,并以热图的形式体现了每只小鼠在科水平分类群(行)相对丰度。c代表HABN在DSS结肠炎小鼠中的体内治疗效果。
这些研究表明,肠道微生物靶向方法对癌症和IBD的治疗潜力。在未来,我们应该将肠道微生物靶向策略用于治疗其他已被证明的、与肠道微生物息息相关的代谢性疾病和精神健康疾病,如肥胖和阿尔茨海默病等。 6 口服纳米药物用于疫苗和免疫耐受
口服疫苗接种策略可产生具有固有和适应性组分的体液和细胞免疫,从而成功产生粘膜和全身保护性免疫。同样,成功的口服耐受策略将在局部肠粘膜区域和全身范围内产生抗原特异性耐受。诱导保护性或耐受性免疫应答需要满足以下几点:(1)将完整且具有活性的抗原成功递送至肠道;(2)穿过粘膜屏障,以及(3)随后抗原呈递细胞(APCs)的免疫调节。在这方面,口服纳米医学为这些挑战提供了潜在的解决方案。纳米粒子内抗原的包裹或包埋可保护药物分子免受pH介导和酶介导的降解,同时防止其在GIT大表面积上稀释。此外,NPs可以通过被动或主动靶向的方式有效地将抗原递送至吞噬APCs,从而刺激抗原特异性细胞和体液应答(表1)。
在开发口服疫苗和耐受策略时,应充分考虑免疫耐受、无能/缺失和保护性免疫应答之间的微妙平衡。首先,应考虑到递送抗原的剂量,以成功进行口服疫苗接种和耐受。通常而言,与传统胃肠外免疫接种相比,口服递送需要更高剂量的抗原来诱导GALT的保护性免疫应答。但过高剂量的抗原会诱导无能/缺失,因此有必要通过口服纳米药物递送足够剂量的抗原进行保护性免疫。另一方面,低剂量重复口服给药可能会触发基于Treg的耐受响应,这也是应该值得注意的事。其次,为了成功进行口服疫苗的接种工作,需要将抗原与免疫刺激佐剂共同递送值靶点,以避免免疫耐受。对于口服耐受,应考虑使用免疫调节药物的共递送抗原。另一个重要因素是靶组织部位。虽然成功的口服疫苗接种需要靶向GALT中M细胞的抗原递送,但固有层中CD103+ DC的抗原递送被证明可诱导免疫耐受。另一方面,将较低剂量的抗原递送至无危险信号的M细胞也可诱导耐受免疫应答。
6.1 口服纳米药物的疫苗应用
目前被许可的口服疫苗都是基于减毒病原微生物和灭活疫苗,其可引起广泛的免疫应答,这些疫苗的特征为血清(IgG)和粘膜(IgA)抗体以及效应T细胞和记忆T细胞。相比之下,考虑到GIT系统的复杂性,目前还没有获得许可的基于亚单位的口服疫苗。为了通过口服途径有效递送抗原,有必要考虑以下几点:1)在转运过程中保护有效载荷免受GIT的恶劣条件;2)在小肠较短的停留时间(3-4 h)内通过粘膜和上皮层向诱导部位的APC递送足量的抗原;3)通过加入佐剂或使用具有固有佐剂特性的疫苗递送载体增强免疫应答(表1)。
携带疫苗抗原的口服纳米药物应通过M细胞和其他肠细胞从肠腔转运至GALT。M细胞可有效内化并将颗粒物(例如细菌或病毒)转运至底层派尔集合淋巴结,因此其被视为口服疫苗设计的理想靶标。M细胞表达多种适合靶向的受体。这些受体包括α-L-岩藻糖残基(与凝集素相互作用)、β1整合素(与cRGD相互作用)、claudin 4(与claudin 4靶向肽相互作用)以及糖蛋白2(与FimH或其他GP2配体相互作用)。例如,在口服免疫实验中,聚集的脂质体能够有效地靶向派尔集合淋巴结中的M细胞,导致具有高抗体滴度的粘膜反应(图9a-e)。虽然M细胞是经口接种的有吸引力的靶标,但M细胞在滤泡相关上皮细胞中的占比较低,其比例尚不到5%。因此,靶向正常肠道上皮细胞是口服疫苗接种的替代策略。已有的研究表明,上皮细胞上表达的多种凝集素(N-乙酰-葡糖胺和唾液酸残基)和PRRs能够跨上皮转运。麦胚凝集素可与N-乙酰基-D-氨基葡萄糖和唾液酸残基结合,Toll样受体(TLR)激动剂已被用作上皮细胞的靶向配体。TLR激动剂,尤其是TLR-2和TLR-4激动剂,能够增强微粒穿过肠腔进入滤泡相关上皮的转运能力(图9f-h)。另一方面,天然聚合物,如靶向CD4的透明质酸和粘膜粘附性壳聚糖,可以与肠上皮结合;因此,基于具有内在靶向能力的天然聚合物的NPs值得进一步被研究。
图9 用于疫苗应用的口服纳米医学。a代表非聚集的脂质体和聚集的脂质体的TEM图像;b代表聚集的脂质体在体内靶向M细胞的能力(箭头所示);c-e分别代表口服携带乙型肝炎表面抗原的脂质体诱导的血清抗体浓度(c),粘膜分泌物中的sIgA浓度(d)以及小鼠脾匀浆中的细胞因子(IL-2和IFN-c)浓度(e);f-g代表FAE中的微球数量(f)和与派尔集合淋巴结FAE中M细胞(红色;g)和上皮内CD11c+ DC(红色;h)相关的微球(绿色)的免疫荧光图像。
对于预防性口服疫苗,有必要采用佐剂来增强免疫应答。产自霍乱弧菌(Vibrio Cholerae)的霍乱毒素和来自产肠毒素大肠杆菌(enterotoxigenic E. coli)的不耐热肠毒素是强效的粘膜佐剂,但这些天然毒素同样也造成了安全性问题。包括TLR激动剂在内的分子佐剂目前以被研究,这些佐剂包括用于TLR4活化的脂多糖或单磷酰脂质A、用于TLR5活化的鞭毛蛋白以及用于TLR9活化的CpG脱氧寡核苷酸。与抗原和TLR激动剂结合的口服NPs能够产生免疫应答。对于具有内在佐剂性的纳米粒子而言,它们可以在不需要佐剂的情况下调节免疫反应。
新冠病毒SARS-CoV-2能够引起肺炎相关呼吸系统疾病(COVID-19),导致了前所未有的国际健康危机。口服免疫作为一种可行的策略,可以预防呼吸系统疾病。例如,口服流感疫苗在II期临床试验中显示了良好的效果,这突出了口服接种在对抗呼吸道病原体中所具有的潜力。此外,口服表达SARS-CoV-2抗原和dsRNA佐剂的腺病毒载体疫苗(称为VXA-CoV-2–1),从1期临床试验(NCT04563702)中获得了可观的初步数据。肠溶包衣用于保护活性成分免受胃酸环境的影响。这使得VXA-CoV-2–1具有很好的耐受性,同时又能触发抗SARS-CoV-2的免疫应答,这种免疫应答表现为抗病毒Spike蛋白的CD8+细胞毒性T细胞应答增加,以及浆细胞和促炎性Th1细胞因子增加。VXA-CoV-2–1也诱导血清和鼻IgA反应。由此可见,应设计、开发和临床试验更多的COVID-19口服纳米医学疫苗。
6.2 用于免疫耐受的口服纳米药
虽然我们的免疫系统可以通过产生全身免疫应答来检测和消除外来病原体,但免疫系统也可以无意中对宿主进行攻击。诸如类风湿性关节炎、多发性硬化、系统性红斑狼疮和1型糖尿病等自身免疫性疾病的根本原因是不适当的自身反应性体液或细胞免疫应答。自身免疫反应对细胞和器官造成严重损伤,有时会产生致命后果。用于治疗的替代蛋白是基因缺陷(如血友病)的常规治疗,但许多患者产生了抗重组生物制剂的中和抗体。例如,凝血因子VIII(F. VIII)和酸性α-葡萄糖苷酶(GAA)已被用作甲型以及乙型血友病和庞贝氏症治疗的替代蛋白,但一部分患者产生了抗重组蛋白的抗体,这限制了对患者的有效治疗。
尽管口服耐受策略在临床前研究中显示了良好的结果,但仍存在一些缺点,诸如可变的pH值、蛋白质降解、需要频繁给药以及较高的成本,这些缺点限制了其广泛应用。由于纳米粒子中掺入的自身抗原的稳定性、自身抗原的调控和延长释放以及纳米粒子的细胞和组织特异性靶向能力,口服纳米粒子可以克服上述的限制(表1)。在一项研究中,与PEG结合的胶原肽通过诱导对胶原的耐受性来抑制胶原诱导的关节炎(图10a-c)。此外,与壳聚糖络合的cDNA已被用于口服免疫耐受。通过cDNA和壳聚糖之间的静电相互作用进行纳米包裹,可保护cDNA不被消化。由于壳聚糖与肠粘液层非特异性结合,口服cDNA-壳聚糖复合物导致其在肠上皮中蓄积,携带F. VIII cDNA的NPs对甲型血友病基因敲除小鼠有效(图10d-g)。在血浆中检测到功能性F.VIII蛋白,相当于2-4%的F. VIII活性,并且在30只小鼠中,有13只小鼠的出血表型得到有效修正(图10e和10f)。另一项研究还发现,负载犬F. VIII cDNA的壳聚糖处理的小鼠血液中未检测到中和F. VIII抗体,表明适当诱导了口服耐受(图10g)。
图10 口服纳米药物诱导免疫耐受。a代表NP/pep(Ⅱ型胶原肽)(A)NP/PEG-pep1(mPEG-SPDP肽)(B)以及NP/PEG-pep2(mPEG-OD肽)(C)的SEM图像;b-c分别代表在DBA/1小鼠派尔集合淋巴结中,基于FACS分析NP/PEG-pep1或NP/PEG-pep2产生IL-4(b)和IL-10(c)的体内T细胞诱导活性;d代表基于定量PCR测定口服壳聚糖NPs后其在生物体内的分布;g-h代表口服FVIII DNA-壳聚糖NPs后,生物体内功能性FVIII蛋白的产生(g)和表型校正(h)活性;i代表通过ELISA介导的血浆FVIIII抗体水平检测。
霍乱毒素B亚基CTB是一种很有前景的口服耐受佐剂。CTB与肠上皮细胞表达的GM1神经节苷脂结合,并通过肌动蛋白依赖性以及ATP依赖性过程促进摄取至固有层。一项基于CTB-GFP融合蛋白来探究口服CTB抗原结合物在生物分布的研究中,学者发现,小鼠口服含有CTB-GFP融合蛋白的叶绿体转基因叶片材料后,可在回肠、肝脏和脾脏中检测到GFP蛋白,这在巨噬细胞和DC内尤为明显。由于树突状细胞在诱导口腔耐受中具有至关重要的作用,CTB可作为将自身抗原递送至固有层树突状细胞的载体。实际上,CTB偶联的自身抗原已被证明能抑制DC活化并诱导Foxp3+ Treg。但CTB和抗原在到达靶点之前,都可能在恶劣的GIT条件下被降解和水解。因此,CTB和抗原通过NPs的稳定共递送将是一个有趣的未来研究方向。在另一个例子中,已经开发了携带CTB的纳米重组痘苗病毒,该重组痘苗病毒是将CTB与胰岛素原基因和谷氨酸脱羧酶的C末端肽融。非肥胖糖尿病小鼠(NOD小鼠)经口给予该系统可降低高血糖和胰岛炎的发病率,与之相对的是,未使用该系统的对照组小鼠则出现高血糖症。此外,基于植物的平台可提供口服耐受的替代方法。口服表达T细胞表位的粉末状水稻种子可诱导变应原特异性口服耐受,改善花粉或螨虫引发的过敏症状。
展望、挑战及未来研究方向
正如本文所讨论的,包括粘膜免疫系统、肠道屏障以及肠道微生物群在内的不同胃肠道(GIT)因子,不仅影响GIT的局部疾病,同时也会对全身不同的系统疾病造成影响。 口服纳米医学提出了新的治疗方法,用以克服GIT的理化和生物学障碍,并选择性地将药物递送到GIT靶位点。因此,口服纳米医学可能为口服疫苗、耐受自身免疫性疾病(如多发性硬化症、类风湿性关节炎)以及IBD、癌症和代谢性疾病(如肥胖、糖尿病)等疾病的治疗带来新的策略。与其他给药途径相比,口服给药途径具有明显的优势,如有限的全身药物暴露、简单的自我给药、良好的患者依从性以及易于在体内分布等。此外,人体共生的肠道菌群与各种疾病的发病机制有关,因此肠道菌群在维持GIT的稳态中起着至关重要的作用。 尽管在过去十年中口服纳米药物取得了进展,但成功的临床转化仍有待解决许多挑战,首先,已有的研究表明,口服疫苗产生的免疫应答差异主要依赖于GIT系统的营养和健康状态。例如,热肠症可导致儿童营养不足和炎症性疾病,从而降低口服免疫的疗效。其次,需要更好地了解剂量和抗原释放动力学,以开发诱导保护性免疫或免疫耐受的有效方法。再者,NPs应在靶位点释放其药物,一旦过早释放药物,则可引起全身吸收介导的副作用。为了防止过早释放,需要在纳米制剂中稳定载药,但这样一来,药物释放情况会随着制剂稳定性的增加而降低。因此,应进一步利用非靶位点和靶位点之间的差异(如酶、pH值、GSH水平和活性氧的不同)进行药物的组织特异性释放。第四,已研究所提及的许多NP系统结构复杂,与之相对应的量产以及质控等问题阻碍了其临床转化。由此可见,口腔纳米医疗系统的设计应尽可能简单。最后,在开发靶向肠道微生物群的口服纳米医学时,应该考虑到个体间肠道菌群可能会有所不同,这取决于人群的年龄、环境暴露、健康状况、遗传、地理和饮食等因素。此外,运动、抗生素使用和手术干预等方式也可改变肠道微生物菌群,这些诸多因素都应成为指定治疗策略时需要考量的额外因素。 除了应该考虑到对GIT的影响,我们认为口服纳米医学应进一步探索用于治疗中枢神经系统(CNS)相关疾病。 肠-脑轴是一个具有密集研究兴趣的新兴领域。由于GIT中存在5亿个神经元支配肠道,其被视为是人体的“第二个大脑”。肠道微生物及其代谢产物影响CNS的发育和体内平衡。肠道细菌通过粘膜和肌肉层迷走神经上表达的受体及其配体(如胆囊收缩素、外源性肽、毒素、5-羟色胺等)与大脑进行通信。诸如阿尔茨海默病和帕金森病等在内的神经退行性疾病,与肠道菌群、其代谢物和淀粉样生物膜共生息息相关。因此,这些可作为药理学的靶点,在未来聚焦于开发能够靶向和正常化肠-脑轴的口服纳米医学,用于CNS疾病的潜在治疗。 综上所述,口服纳米医学为调节肠道菌群和微生物代谢产物提供了一种可行的策略,并为各种生物医学应用提供了有前景的治疗平台。
不感兴趣
看过了
取消

人点赞

人收藏

打赏
不感兴趣
看过了
取消
©2012-2023 北京华媒康讯信息技术股份有限公司 All Rights Reserved. 注册地址:北京 联系电话:010-82736610
广播电视节目制作经营许可证 —(京)字第 17437号 京海食药监械经营备20200522号
京ICP备12011723号 京ICP证150092号
 京公网安备 11010802020745号
工商备案公示信息
互联网药品信息服务资格证书((京)-非经营性-2020-0015)
京公网安备 11010802020745号
工商备案公示信息
互联网药品信息服务资格证书((京)-非经营性-2020-0015)






您已认证成功,可享专属会员优惠,买1年送3个月!
开通会员,资料、课程、直播、报告等海量内容免费看!

打赏金额
认可我就打赏我~
1元 5元 10元 20元 50元 其它
打赏作者
认可我就打赏我~
扫描二维码
立即打赏给Ta吧!
温馨提示:仅支持微信支付!
已收到您的咨询诉求 我们会尽快联系您











































 010-82736610
010-82736610
 股票代码: 872612
股票代码: 872612




