
许多研究表明,LAB菌株可能会产生一种抗癌物质,该物质是细胞壁的组成部分。革兰氏阳性菌(包括乳酸菌)细胞壁的主要化合物是肽聚糖(PG)。
导读
乳酸菌(Lactic acid bacteria,LAB)是一种天然存在于发酵食品中的革兰氏阳性微生物,多年来一直受到研究人员的关注。LAB是具有多种功能和活性的生物活性化合物的有效来源,尽管尚未完全了解。LAB的代谢产物,如短链脂肪酸、胞外多糖和细菌素等都具有很好的抗癌潜力。
对LAB生物活性代谢物与免疫机制相互作用的研究表明,这些物质可以发挥强大的免疫调节作用,这也解释了其巨大的治疗潜力。LAB的抗癌活性在体外和动物模型中都得到了证实,可对抗各种恶性肿瘤的癌细胞。LAB通过多种机制抑制肿瘤生长,如抗增殖活性、诱导凋亡、细胞周期阻滞,以及抗诱变、抗血管生成和抗炎作用。
本综述旨在总结有关LAB抗癌活性的最新数据,侧重于最有前景的具有潜在临床应用的生物活性化合物。
论文ID
原名:Anticancer activity of lactic acid bacteria
译名:乳酸菌的抗癌活性
期刊:Seminars in Cancer Biology
IF:15.707
发表时间:2022.1.4
通讯作者:Katarzyna Garbacz
通讯作者单位:波兰格但斯克医科大学医学院
DOI号:10.1016/j.semcancer.2021.12.013
综述目录
1 前言
2 乳酸菌特性
3 乳酸菌是肠道微生物群的组成部分
4 肠道中乳酸菌的抗癌活性
5 乳酸菌抗癌活性的机制
6 LAB的抗癌化合物
6.1 细菌素
6.1.1 乳酸链球菌肽 6.1.2 Plantaricin A 6.1.3 片球菌素 6.1.4 Biovicin HC5
6.2 酶类
6.2.1 精氨酸脱亚胺酶
6.3 细菌细胞壁
6.3.1 肽聚糖 6.3.2 表层 6.3.3 胞外多糖
6.4 短链脂肪酸
7 结论
主要内容
1 前言
多年来,恶性肿瘤一直是全世界死亡的主要原因。传统的治疗方法,如化疗和放疗,虽然能有效控制癌症的生长和扩散,但也产生多种不良反应。因此,肿瘤药理学研究的一个重要目标是确定新的抗肿瘤治疗方案,与传统方法一样有效,但副作用更少。具有抗癌潜力的细菌物质,如乳酸菌(LAB)代谢产物,似乎是一种有前景的合成抗肿瘤药物的替代品。
许多微生物化合物的潜在临床应用是最近才发现的。LAB的生物活性代谢物在体外和动物研究中都产生了很好的结果,其中一些已经成为临床试验的主题。本篇综述旨在总结有关LAB抗癌活性的最新数据,特别强调最有前景的具有潜在临床应用的生物活性化合物。
2 乳酸菌特性
乳酸菌(LAB)是一个同质类群,其共同特征是碳水化合物发酵的主要产物乳酸的合成。LAB为革兰氏阳性、过氧化氢酶阴性杆菌或球菌,主要来自乳杆菌目(Lactobacillales)。一般来说,LAB不产生孢子、不流动、厌氧或微好氧,它们是嗜酸性菌,这使它们能够在pH值(4.5-7.0)相对较低的环境中生存和繁殖。随着环境酸度的增加,LAB对环境酸度的敏感性降低,主要由精氨酸代谢酶、精氨酸脱氨酶和精氨酸酶决定。LAB可以在各种环境中很好地工作,因为它们可以在较宽的温度范围内增殖(10-45℃),盐度高达6.5% NaCl。
LAB的另一个特征是其基因组中G+C碱基对的数量较少。 根据发酵产物的不同,LAB可分为同质发酵型和异质发酵型。前者发酵的主要产物是乳酸,而后者则合成乳酸和其他产物,如乙酸、乙醇和二氧化碳。 按照目前的分类体系,LAB属于厚壁菌门(Firmicutes)、芽孢杆菌纲(Bacilli)和乳杆菌目(Lactobacillales)。根据它们的细胞形态、葡萄糖发酵的类型、生长温度和发酵糖的范围进行分类。
LAB菌属主要有乳杆菌属(Lactobacillus)、明串珠菌属(Leuconostoc)、片球菌属(Pediococcus)、乳球菌属(Lactococcus)和链球菌属(Streptococcus),其他菌属的生物学重要性较低,如Aerococcus、Alloiococcus、Carnobacterium、Dolosigranulum、肠球菌属(Enterococcus)、Oenococcus、Tetragenococcus、Vagococcus和魏斯氏菌(Weissella)。Lactobacillus是上述列出的属中最大的,已描述的种类超过100种。Bifidobacterium spp.虽然不属于乳杆菌目,但也被归类为LAB,因为它们能够合成乳酸作为发酵产物(图1)。
LAB是普遍存在的微生物,通常在牛奶、乳制品和植物衍生物质中发现。它们存在于发酵乳制品、蔬菜、肉类和鱼类产品、酵母、青贮和发酵饮料中。此外,LAB还在人类和动物中定居,可从女性的口腔、回肠、结肠和阴道中分离出来。除哺乳动物外,LAB也普遍存在于鱼类、两栖动物、爬行动物和鸟类的胃肠道中。
LAB属于食品工业和生物技术中最重要的微生物群,它们通常被用作乳制品和发酵植物衍生产物生产的发酵剂。它们不仅有助于改善许多产品的口感和质感,而且由于它们能够抑制与食物腐烂有关的细菌,其还被用于保存食物。LAB的抑制特性主要与乳酸的合成和环境pH值的降低有关。
然而,LAB还可以产生其他抑制细菌生长的物质,如细菌素、过氧化氢和二乙酰。 由于LAB对健康有益,一些乳酸菌被用作益生菌,并被视为GRAS(通常被认为是安全的)状态,这意味着如果使用得当,它们是安全的。除了一般的非致病性外,益生菌LAB还需要满足许多其他的选择标准、功能和技术要求。这些标准包括来自人类微生物群、严格定义的属和种分类、抗胃液和胆盐降解性、能够粘附上皮细胞的表面特性、合成抗菌物质的能力、以及对典型细菌病原体的拮抗作用。此外,益生菌LAB应被宿主的免疫系统耐受良好,不应诱导产生抗体。
在授权特定LAB作为人类或动物的益生菌使用之前,需详细规定并仔细验证这些要求。最有名的LAB菌株属于乳杆菌属(Lactobacillus)、乳球菌属(Lactococcus)、肠球菌属(Enterococcus)、链球菌属(Streptococcus)、片球菌属(Pediococcus)和明串珠菌属(Leuconostoc)。
图1 乳酸菌菌属
3 乳酸菌是肠道微生物群的组成部分
LAB是肠道微生物群的重要组成部分。成人的肠道是一个复杂的生态系统,每1克肠道内容物中含有大约1000种不同种类的1012个微生物。细菌定植在出生后立即开始,并持续数年。肠道微生物群的最终组成取决于多种因素,具体到每个个体,由多个细菌群体组成,其中包括LAB。
肠道内定植的LAB有巨大的代谢和酶促潜能;它们参与许多外源性和内源性化合物的代谢,合成具有多种活性的产物。LAB形成肠道微生物群的主要特征包括粘附在粘膜上,在不构成感染风险的情况下在肠隐窝中定植,以及一旦种群被破坏(例如由于抗生素治疗)后自我再生的潜力。肠道微生物群的稳定性可能是由于肠道免疫系统产生耐受所致。 生理性肠道菌群是消化道正常功能所必需的。
现有证据表明,肠道菌群对免疫系统的正常功能和营养物质的正常消化也至关重要。肠道微生物群的主要作用是防止感染和条件性微生物的过度生长。肠道细菌调节肠细胞的生长、分化和再生,具有免疫调节特性,影响消化道的运动功能。
此外,肠道细菌还能降解食物中的毒素和致癌物,合成微量元素,代谢未被人体系统消化的食物残渣,参与电解质和矿物质的吸收。 生理性肠道菌群的功能与其免疫调节活性有关。肠道细菌可增强体液反应,激活非特异性免疫机制,调节对潜在病原体的免疫反应,并减轻超敏反应。肠道微生物群对免疫系统的成熟及其活性的维持起着至关重要的作用。肠道相关淋巴组织(GALT)出生后成熟。
在这一过程中,细菌菌群在肠道定植的刺激起着至关重要的作用,特别是它们与肠上皮细胞和树突状细胞的相互作用。LAB对免疫的有益作用主要与肠上皮细胞和树突状细胞表面模式识别受体(PRRs),特别是toll样受体(TLR)的特异性激活有关。这些受体识别LAB特定的分子模式,即所谓的微生物相关分子模式(MAMPs),导致细胞内信号的多重级联激活(特别是NF-kB转录因子),并合成内源性抗菌因子、防御素、促炎细胞因子和趋化因子。这最终导致肠道屏障开放机制的激活,并限制肠道内微生物的存在。然而,与此同时,LAB的一些代谢产物,如自由基和脂多糖,可阻止NF-kB抑制剂(IkB)的泛素化和降解,从而限制该途径的激活。此外,LAB的MAMP对TLR的激活导致PPAR-g的诱导,限制了细胞核内NF-kB的作用,促进了防御素的合成,从而阻止了炎症的发生。
4 肠道中乳酸菌的抗癌活性
早在20世纪70年代末,就有人证明,食用含有嗜酸乳杆菌(Lactobacillus acidophilus)的发酵牛奶会导致老鼠肠道中这些细菌数量的增加,这反过来又会导致腐烂细菌数量的减少,如大肠菌群参与肿瘤促进因子和前致癌物的合成。
随后,Moore等人对肠道微生物群的分析表明,某些LAB菌株的存在与较低的癌症风险之间存在联系。许多最近的动物研究结果表明,LAB作为益生菌使用可能在控制肠道癌的发生中发挥重要作用。据称具有抗癌活性的益生菌包括各种LAB制剂和发酵乳制品。日本的一项队列研究显示,饮用酸奶可以显著降低结直肠癌死亡率,该研究的参与者超过10万人。
此外,在瑞典和意大利进行的大型(约4.6万名参与者)前瞻性队列研究分别持续了7年和12年,证明了结直肠癌的发病率和乳制品消费(包括发酵乳制品)之间存在明显的负相关关系。 LAB的抗癌活性研究一般涉及化学诱导的小鼠类致癌模型;根据实验的持续时间,研究涉及到异常隐窝病灶(ACFs),其被认为是结直肠癌(CRC)的前体病变。许多研究检查了LAB作为益生菌使用对CRC和前体病变(ACFs)发展的影响。
分析表明,益生菌对ACFs和CRC均有明显的抑制作用。总体上,检测了54种益生菌制剂,其中33种在ACF模型中,26种在CRC模型中。共有22/33和16/26的受检制剂分别被证明能阻止ACF和CRC的发展。在其他一些研究中,含有益生菌嗜热链球菌(Streptococcus thermophilus)、保加利亚乳杆菌(Lactobacillus bulgaricus)、青春双歧杆菌(Bifidobacterium adolescentis)、干酪乳杆菌(Lactobacillus casei)、长双歧杆菌(Bifidobacterium longum)、乳酸乳球菌(Lactococcus lactis)和屎肠球菌(Enterococcus faecium)菌株的酸奶培养物与Lactobacillus helveticus spp. jugurti相结合,对ACFs或CRC的发生没有影响。
有些菌株被证明只有在与益生元(以共生的形式)结合时才有效。其他涉及动物模型的研究也表明,补充LAB可预防CRC。 在最近的一篇报道中,Wang等人研究了Bifidobacterium bifidum CGMCC 15068对偶氮甲烷(AOM)/葡聚糖硫酸钠(DSS)诱导的结肠炎相关CRC(CAC)小鼠模型的有益作用。AOM(10 mg/kg)腹腔注射和DSS在饮用水中给药3周,两个连续周期之间有14天的恢复期。
两歧双歧杆菌CGMCC 15068对CAC小鼠模型的致癌性有一定的抑制作用。研究结果支持了两歧双歧杆菌CGMCC 15068通过调节肠道菌群失调和代谢方面在肠道健康中的有益作用。因此,益生菌两歧双歧杆菌(B. bifidum)菌株似乎是一种预防CRC的有前景的策略。
Ishikawa等人分析了一种益生菌干酪乳杆菌(L. casei)菌株对398例切除至少两个息肉或早期结直肠恶性肿瘤患者的结直肠癌发展的影响。经过4年的随访,干酪乳杆菌菌株补充组中中度或高度异型性肿瘤的发生率显著低于对照组。然而,在同时补充膳食纤维的患者组中未观察到益生菌的有益作用。 在随机双盲安慰剂对照研究中,息肉或结肠恶性肿瘤切除后的患者补充益生元和益生菌Bifidobacterium lactis Bb12和Lactobacillus delbrueckii subsp. rhamnosus GG菌株12周。
补充组病人的上皮细胞被证明不容易受到肠道中存在的细胞毒性和基因毒性因子的有害影响。此外,补充益生菌有助于增加有益肠道微生物的数量。在健康志愿者参与的临床实验研究中,益生菌的抗癌活性也得到了证实。在一项随机双盲安慰剂对照研究中,健康志愿者接受了含有活L. rhamnosus LC705细胞的胶囊或安慰剂胶囊,结果显示有益肠道细菌增加,粪便β-葡萄糖苷酶、β-葡萄糖醛酸酶和脲酶活性显著降低。 补充LAB可使结肠癌患者的肠道微生物群结构发生有益变化,并刺激患者的免疫系统。
Gianotti等人进行了一项随机安慰剂对照临床试验,证实了两种益生菌LAB菌株——Bifidobacterium longum BB536和Lactobacillus johnsonii La1在该患者组中的有益作用。同样,一项结肠切除术后患者接受Lactobacillus plantarum OCGMCC 1258、Lactobacillus acidophilus LA-11和Bifidobacterium longum BL-88混合益生菌的研究改善了肠道屏障的完整性,并降低了患者围手术期感染相关并发症的风险。
5 乳酸菌抗癌活性的机制
LAB的抗癌作用似乎是多方面的,其一些潜在机制尚未完全了解(图2)。关于LAB抑制肠道癌变,假设将这些细菌与食物一起摄入可增加肠道有益菌群的数量,从而抑制参与肿瘤促进因子和前致癌物合成的致病微生物的生长和活性。选择适当的乳酸菌可以争夺营养物质,合成对病原菌有拮抗作用的物质(细菌素、乳酸)。
此外,LAB刺激与致病微生物抗原相互作用的抗体的产生,并阻断肠道受体,从而显著减少肠道内病原菌的存在,这导致粪便酶、β-葡萄糖苷酶、β-葡萄糖醛酸酶、硝酸还原酶和硝基还原酶活性下降。另一个被认为参与LAB抗癌活性的重要机制是短链脂肪酸(SCFAs)的合成,主要是乙酸、丙酸和丁酸。短链脂肪酸主要是低聚果糖发酵的产物,有助于降低肠道pH值。这一机制对维持肠道内环境稳态和正常生理过程至关重要。较低的pH限制了条件性菌群的生长,并调节肠道内的酶活性。
一旦合成,SCFAs被各种细胞和组织吸收,影响其生长、增殖和凋亡。 许多体外和体内研究表明,LAB可以降低肠道中粪便酶的活性,主要是β葡萄糖醛酸酶、硝酸还原酶和硝基还原酶。肠道长期暴露于外源性物质,如药物、食品防腐剂和环境污染物。这些外源性物质中只有一部分会以其原生形式从体内清除。大多数外源性物质在羟基化和结合过程中发生变化,主要发生在肝脏;这些过程改变了它们的生物活性和水溶性,促进了它们的消除。粪便中的酶干扰这些过程,参与致癌物、基因毒素和肿瘤启动子的合成和激活。
绝大多数外源性物质通过葡萄糖醛酸化在肝脏中进行结合和解毒,所生成的葡萄糖醛酸与胆汁一起释放到肠道。由于其底物特异性较低,存在于肝脏的细菌β-葡萄糖醛酸酶可以水解这些代谢物,释放致癌苷元。 对结直肠息肉或结直肠癌患者的研究证实了粪便中胆汁酸(特别是去氧胆酸)浓度与结直肠恶性肿瘤发病率之间的联系。
肠道细菌参与胆汁酸代谢,导致致癌性次级胆汁酸浓度增加。次级胆汁酸影响肠细胞的坏死、增生和致癌活性,诱导DNA损伤和细胞凋亡。也有人推测,可溶性胆汁酸可能对肠细胞产生细胞毒性作用,导致其过度增殖。结肠癌患者给药益生菌嗜酸乳杆菌(L. acidophilus)菌株,可降低粪便中可溶性胆汁盐的浓度。
结肠息肉患者补充嗜酸乳杆菌和Bifidobacterium bifidum菌株会导致大肠pH值降低并减少癌细胞的增殖。 LAB在结合和/或降解潜在致癌物和合成抗癌物质的能力方面各不相同。早在20世纪80年代就有研究表明LAB可以降解亚硝胺,这种诱变化合物通常存在于腌肉和干鱼中。
一项体外研究表明,LAB可以结合杂环胺和其他食物源诱变剂,降低其活性。在喂食含有LAB饮食的小鼠的各种组织中,一种强诱变化合物的同化减少,这种化合物通常存在于油炸肉和鱼中。反过来,食用大量油炸肉类的受试者服用干酪乳杆菌(L. casei)可降低其尿液和粪便的致突变性。这种效应似乎与益生菌结合并消除肠道中的诱变因子有关。
LAB多糖的抗癌活性与刺激免疫细胞(主要是T、B淋巴细胞、巨噬细胞和NK细胞)释放白细胞介素有关。L. plantarum 299v菌株具有减轻肠道黏膜炎症并促进其再生的作用。根据Rask等人的研究,发现补充L. plantarum 299v可增加CD8+ T细胞上激活标志物CD25和CD4+细胞上记忆标志物CD45RO的表达。即使这些T淋巴细胞表面标志物活性的变化不是由L. plantarum直接引起的,这种LAB也可以通过抗原呈递细胞(APC)间接刺激淋巴细胞。
图2 乳酸菌的抗肿瘤作用
6 LAB的抗癌化合物
LAB具有高代谢活性和产生广谱抗菌化合物的特性;其中一些最近已经在抗癌活性方面进行了研究。LAB合成的化合物通常具有较低的细胞毒性且产生的副作用很少。因此,它们似乎是合成抗癌剂的一个很好替代品。
6.1 细菌素
LAB产生的细菌素是一种在核糖体中合成的多肽和蛋白质,具有不同的结构、光谱和作用机制。通常,细菌素是由30-60个氨基酸组成的带正电荷的小分子物质,主要对革兰氏阳性细菌有效,其活性主要与细菌细胞质膜内膜电位的破坏有关,导致离子不受控制的泄漏和细胞死亡。
类似的机制也决定了细菌素的抗癌活性(表1)。 LAB产生的细菌素可分为三类。I类是分子质量小于5 kDa的羊毛硫抗生素;由于翻译后修饰,这些化合物获得了独特的氨基酸,如羊毛硫氨酸、3-甲基羊毛硫氨酸、脱氢丙氨酸和脱氢丁氨酸。I类细菌素的两个代表性化合物是乳酸链球菌肽和乳酸菌素。
II类细菌素是小分子(<10 kDa)、热稳定、不可修饰的翻译后肽;它们可分为两个亚类,IIa和IIb。亚类IIa包括具有强生物活性的片球菌素(pediocins),其中最有名的化合物是pediocin PA1和leucocin A。亚类IIb细菌素是需要两个互补肽协同活性才能显示抗菌活性的化合物;该亚类的两个代表性化合物是plantaricin A和enterocin X。III类细菌素是分子质量超过30 kDa的大分子且不耐热的蛋白质,例如helveticin J。
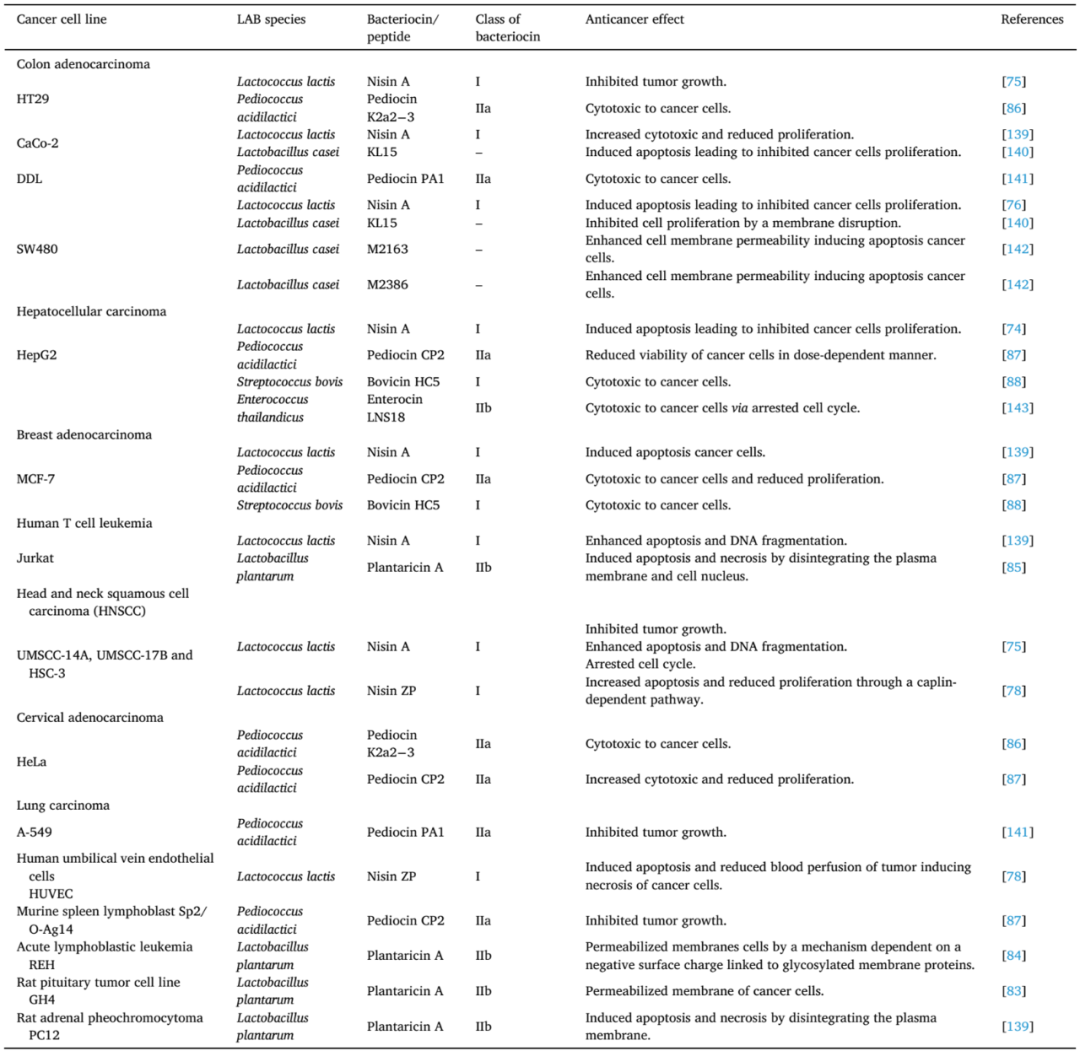
表1 乳酸菌(LAB)产生的细菌素和多肽对癌细胞系的活性
6.1.1 乳酸链球菌肽
乳酸链球菌肽A(Nisin A)是最常用和最著名的LAB细菌素之一。这种化合物是近一个世纪前(1928年)由Rogers和Whittier发现的,从发酵的牛奶中分离出来。乳酸链球菌肽A是由Lactococcus lactis合成的由34个氨基酸组成的环状肽。它是一种转座子和质粒编码的翻译后修饰的AMP,很容易在L. lactis菌株之间转移。乳酸链球菌肽A分子质量为3.49 kDa,具有广谱抗菌活性,对革兰氏阳性菌、革兰氏阴性菌和耐酸分枝杆菌均有抗菌作用。
乳酸链球菌肽A在非常低的浓度和低pH值下具有杀菌作用[19 O’Suillvan],它被认为对人类和动物都是安全的,因此被授予GRAS。由于上述特性,乳酸链球菌肽A作为食品防腐剂(E234)已在全世界近50个国家(包括欧盟成员国和美国)使用了50多年(自1969年以来)。
体外和体内研究结果表明乳酸链球菌肽A具有抗癌活性。Joo等人发现乳酸链球菌肽A抑制了三种不同人类细胞系(即UMSCC-14A、UMSCC-17B和HSC-3)中头颈部鳞状细胞癌(HNSCC)的增殖。在小鼠模型中,乳酸链球菌肽A也被证明可以防止体内HNSCC的发展。随着细菌素浓度的增加(从5 μg/mL增加到80 μg/mL),癌细胞凋亡和DNA断裂均增强。
阳离子转运调节剂同源物1(CHAC1)也可部分诱导癌细胞凋亡,导致钙离子流入细胞,细胞周期停滞。此外,Ahmadi等人发现乳酸链球菌肽A诱导CRC细胞(SW480)凋亡,具有显著的抗增殖作用,并导致mRNA和蛋白水平上凋亡指数(bax/bcl-2比值)升高。
最近的证据表明,乳酸链球菌肽A不是这个基团中唯一具有抗癌特性的化合物。除乳酸链球菌肽A外,该AMP的其他几种天然变体由L. lactis(nisins Z、F和Q)和其他细菌(如Streptococcus(nisins H、U、U2和P))合成。最近,Kamarajan等人在体外和体内都证实了一种天然的乳酸链球菌肽变体nisin ZP(3.47 kDa),与nisins A具有类似的抗癌潜力。
用浓度不断增加的nisin ZP处理的HNSCC(UM-SCC-17B和HSC-3)细胞通过Caplin依赖性途径显示细胞凋亡水平增加和增殖减少。Nisin ZP还可诱导人脐静脉内皮细胞(HUVEC)凋亡并降低肿瘤的血液灌注,最终导致癌细胞坏死。在小鼠研究中,nisin ZP可抑制肿瘤生长并延长动物的生存时间。
此外,经nisin ZP处理的小鼠解剖器官无炎症、纤维化和坏死迹象。 乳酸链球菌肽作为有前景的抗癌药物的另一个理由是,它们很容易被修饰;目前正在进行的广泛研究旨在获得具有比上述更强抗癌活性的乳酸链球菌肽的新变体。
6.1.2 Plantaricin A
Lactobacillus plantarum C11产生的Plantaricin A具有双重作用,既作为影响基因调控的信息素,又作为抗菌肽。 Plantaricin A的抗菌谱相对较窄,主要包括多种乳杆菌,如L. viridescens、L. sakei和L. casei。Plantaricin A最初被鉴定为细菌素,属于IIb亚类细菌素,不经历翻译后修饰。Plantaricin A分子具有两亲性螺旋结构,分子量为2.98 kDa,其作用的主要机制,无论是针对癌细胞还是细菌都涉及到增加质膜的渗透性,并导致其分解。
在研究Plantaricin A抗癌活性的首项研究中,Sand等人证明了这种化合物能渗透大鼠垂体癌细胞,但不能渗透正常细胞,并在外膜小叶和内膜小叶之间分化。6年后,该团队报告Plantaricin A通过一种依赖于与糖基化膜蛋白连接的表面负电荷的机制渗透真核细胞的细胞膜。
反过来,Zhao等人证明了人工合成的Plantaricin A对人白血病T细胞的体外细胞毒性(Jurkat)。用25 μM剂量的细菌素处理可导致细胞活力下降75%。人工合成的Plantaricin A通过分解质膜和细胞核来诱导Jurkat细胞系凋亡和坏死。此外,这种细菌素治疗与癌细胞内caspase-3浓度的增加有关。
6.1.3 片球菌素
片球菌素(Pediocins)是一组低分子量的IIa亚类细菌素,由Pediococcus spp.合成。Pediocin PA-1由P. acidilactici K2a2-3合成,对宫颈癌细胞(HeLa)和人结肠癌细胞(HT29)具有细胞毒性。根据Villarante等人的研究,PA-1抑制了53.7%的HT29细胞,但仅抑制了15.6%的HeLa细胞。
Pediocin CP-2由P. acidilactici MTCC 5101菌株合成,对少数癌细胞具有高于PA-1的细胞毒性。本文包括CP2的天然形式和重组形式(rec-pediocin)。根据Kumar等人的研究,小鼠脾淋巴母细胞系Sp2/O-Ag14对rec-pediocin CP2最敏感。25 μg/mL的rec-pediocin可使细胞存活率下降到0%,而相同剂量的native pediocin可使Sp2/O-Ag14细胞的存活率保持在26.7%。
其他被分析的细胞系,HeLa、人肝癌细胞(HepG2)和人乳腺癌细胞(MCF-7),对native和rec-pediocin表现出不同的敏感性。25 μg/mL的rec-pediocin和native pediocin CP2对HepG2细胞系的存活率分别为5.5%和1.2%,MCF-7细胞系的存活率分别为2.1%和10.7%。HeLa细胞对rec-pediocin的敏感性低于该研究中包括的其他细胞系。
6.1.4 Biovicin HC5
Biovicin HC5是一种小分子(2.4 kDa)细菌素,属于羊毛硫细菌素类,由Streptococcus bovis HC5菌株产生。广泛的Biovicin HC5生物活性不仅包括多种革兰氏阳性细菌;该化合物还显示出对癌细胞系的体外细胞毒性。Bovicin HC5对人乳腺癌MCF-7细胞系和肝癌HepG2细胞系均有抑制作用。根据Paiva的研究,Biovicin HC5对这两种细胞系的最大半数抑制浓度(IC50)分别为279.4 μM和289.3 μM。
6.2 酶类
6.2.1 精氨酸脱亚胺酶
精氨酸脱亚胺酶(ADI)是体内一种将精氨酸水解成瓜氨酸和铵的酶。ADI可能由一些细菌分泌,包括LAB物种,如粪肠球菌(Enterococcus faecalis)、血链球菌(Streptococcus sanguis)、酒酒球菌(Oenococcus oeni)、Lactococcus lactis subsp. lactis、Lactobacillus sakei、植物乳杆菌(Lactobacillus plantarum)和L. sanfranciscensis。
精氨酸脱亚胺酶可能通过消耗细胞外氨基酸(如精氨酸)的供应来抑制某些肿瘤细胞系的生长。补充精氨酸对细胞生长至关重要。在正常的人体细胞和组织中,瓜氨酸可以通过尿素循环酶精氨酸琥珀酸合成酶(ASS)和精氨酸代琥珀酸裂解酶(AL)生成精氨酸。然而,一些人肿瘤细胞不表达ASS,因此精氨酸营养不良。
因此,精氨酸脱亚胺酶是治疗精氨酸营养不良性肿瘤的潜在癌症治疗剂。精氨酸脱亚胺酶已被证明可以抑制精氨酸营养不良性肿瘤的生长,如黑色素瘤和肝细胞癌(HCC)。ADI还可诱导人白血病细胞周期阻滞和凋亡。此外,还报道了ADI活性对内皮细胞的抗增殖和抗血管生成作用。
在最近的研究中,聚乙二醇修饰的精氨酸脱亚胺酶(ADI-PEG20)已被使用。ADI-PEG20的抗原性降低,血清稳定性提高(半衰期)。聚乙二醇化ADI似乎是一种很有前景的HCC药物,已被用于治疗不能切除和转移性肝细胞癌的II期临床研究。此外,Kim等人发现ADI-PEG20通过诱导CWR22Rv1癌细胞自噬和caspase非依赖性凋亡来治疗前列腺癌的疗效。此外,在四种胶质母细胞瘤细胞系(HROG02、HROG05、HROG10和HROG17)中也观察到了与ADI细胞凋亡无关的抗癌作用。
6.3 细菌细胞壁
6.3.1 肽聚糖
许多研究表明,LAB菌株可能会产生一种抗癌物质,该物质是细胞壁的组成部分。革兰氏阳性菌(包括乳酸菌)细胞壁的主要化合物是肽聚糖(PG)。肽聚糖不仅保护细菌细胞的完整性和细胞形状,而且调节免疫反应,刺激肿瘤坏死因子、干扰素和白细胞介素(IL-1、IL-6、IL-8、IL-12)的产生。
此外,有报道称LAB菌株细胞壁部位的肽聚糖在某些肿瘤细胞系中具有细胞毒性,在体外对不同的肿瘤细胞系有显著的抗增殖作用。Kim等人发现了Lactococcus lactis ssp. lactis的细胞质部分对人胃癌细胞SNU-1具有较强的抗增殖活性,该研究表明L. lactis对SNU-1的抗增殖活性与诱导细胞凋亡有关。
在进一步的研究中,Matsumoto等人指出,Lactobacillus casei Shirota菌株中的完整细胞和细胞壁衍生的肽聚糖多糖(PSPG)均抑制IL-6的产生。他们在动物模型中证实了肽聚糖复合物在结肠炎相关癌症(CAC)抗炎作用中的关键作用。最近,Tian等人报道了从Lactobacillus paracasei subsp. paracasei X12菌株中提取的肽聚糖可诱导人结肠癌细胞系(HT-29)凋亡。类似地,Wang等人从Lactobacillus paracasei subsp. paracasei M5菌株中发现了全肽聚糖(WPG),其可抑制结肠癌HT-29细胞增殖,诱导细胞凋亡。
肽聚糖通过上调促凋亡基因、下调抗凋亡基因和促进线粒体中细胞色素C向胞质释放来介导其凋亡特性。He等人还比较了硒化肽聚糖(Se-PG)与非修饰肽聚糖在HT-29细胞中的抗肿瘤活性。结果表明,Se-PG比天然肽聚糖具有更强的抗肿瘤活性,且具有剂量依赖性。这些结果表明硒化修饰可增强PG的抗肿瘤活性,Se-PG可发挥其潜在的抗肿瘤活性。
6.3.2 表层
表层(S层)是许多乳酸菌的最外层细胞表面。在以下菌株中证实了存在S层:L. acidophilus、L. amylovorus、L. brevis、L. crispatus、L. gallinarum、L. gasseri、L. helvetius和L. johnsonii。S层是由蛋白质或糖蛋白组成的晶体状二维细菌结构。表层的蛋白质占细菌细胞总蛋白质含量的10-20%。
S层蛋白的作用尚未完全了解,但最近的研究表明,它们可能在不同的免疫反应中发挥重要作用。最近,Suzuki等人比较了从8种不同的乳杆菌菌株中分离的S层蛋白的免疫学特性。结果表明,不同种类的乳杆菌S层蛋白具有不同的免疫特性,Lactobacillus amylolyticusNRIC 0558 T蛋白的N端区域具有较高的免疫原性。
在本研究中,Zhang等人研究了Lactobacillus acidophilus CICC 6074的S层蛋白对人结肠癌HT-29细胞的影响。S层蛋白对HT-29细胞具有抗增殖和细胞毒性作用。它通过下调细胞周期素依赖性激酶和细胞周期素B的表达,将细胞周期阻滞在G1期,从而抑制肿瘤细胞的增殖。观察到细胞凋亡的变化,包括染色质凝结、核断裂等。此外,研究表明S层蛋白可能通过线粒体途径诱导HT-29细胞凋亡。L. acidophilus CICC 6074的S层蛋白可能是一种潜在的抗肿瘤药物。
6.3.3 胞外多糖
LAB可以合成大量的胞外多糖(EPS)。胞外多糖是一种高分子量、长链线性生物聚合物,通常具有侧链,由α-和β-糖苷键连接的重复碳水化合物单元组成。EPS可连接到LAB细胞表面,形成胶囊(荚膜多糖,CPS)或以粘液的形式释放到细胞外(粘液多糖)。
EPS在细菌细胞中发挥多种功能;它们保护细菌免受不利的环境和生物因素(噬菌体、宿主的免疫反应)的影响,并促进在新环境生态位的定植。 根据其化学成分,LAB胞外多糖可分为两类,同多糖(HoPS)和杂多糖(HePS)。同多糖由单一类型的单糖、D-吡喃葡萄糖或D-呋喃果糖组成。杂多糖由含有各种单糖或其衍生物的重复亚基构成,它们可以含有侧链或形成含有3-8个糖残基的线性结构。 HoPS由细胞外转移酶合成,被称为聚糖蔗糖酶,它利用蔗糖作为葡萄糖或果糖的来源。
LAB可以合成多种不同的HoPS,其中最有名的是由重复的半乳糖单元构成的α-D-葡聚糖(右旋糖酐、mutan、alternan、reuteran)、β-D-葡聚糖、果聚糖(左旋、菊粉型)和多聚半乳糖。 右旋糖酐由葡萄糖残基与α-1,6糖苷键连接而成,不同程度的分支是LAB物种的一个特征。右旋糖酐可由多种Leuconostoc spp.、Lactobacillus spp.和Streptococcus spp.合成,最早被证明能产生右旋糖酐的微生物是van Tieghem在1878年描述的肠系膜Leuconostoc mesenteroides。L. mesenteroides NRRL B-512F产生的右旋糖酐具有线性结构,95%的葡聚糖由D-葡萄糖残基与α-1,6-糖苷键连接而成。葡聚糖是最早也是最容易理解的HoPS之一,在食品、化妆品和医药工业中有许多应用。
Alternan是一种高水溶性多糖,由D-葡萄糖残基与交替的α-1,6和α-1,3-糖苷键连接而成,它是由一些肠系膜Leuconostoc mesenteroides菌株(NRRL B-1355、NRRL B-1501和NRRL B-1498)以蔗糖为原料合成的。 与上面提到的其他HoPS不同,Mutan不溶于水;这种多糖由主要与α-1,3-糖苷键连接的D-葡萄糖残基组成。Streptococcus mutans是Mutan最主要的来源,它被认为是蛀牙的原因之一。
Mutan被认为是促进口腔链球菌粘附在牙齿表面的主要因素,从而导致牙菌斑的形成。最近的证据表明,Leuconostoc spp.和Lactobacillus spp.也可以产生Mutan,Reuteran主要由Lactobacillus reuteri合成,含有α-1,4-和α-1,6-糖苷键,是水溶性的。
LAB胞外多糖的抗癌活性与其抗增殖和促凋亡的特性有关。研究结果表明,在不同的癌细胞系中,胞外多糖具有一定的抗增殖作用,且具有剂量和时间依赖性。研究最广泛的细胞系来自肠道、肝脏和乳腺恶性肿瘤。根据许多作者的研究,LAB制剂在肠道癌细胞系中具有最强的抗增殖作用,这意味着它们在这些恶性肿瘤中的治疗潜力可能是最高的。这可能是由于EPS容易与肠道癌细胞表面受体结合的结果,也可能是LAB在人类肠道的自然发生。LAB胞外多糖诱导的细胞凋亡特征性变化包括细胞核解体、染色质凝聚、细胞质空泡化和浓缩。
已知LAB诱导的细胞凋亡的主要机制是基于caspases控制的外源和内源途径的激活,caspases是天冬氨酸特异性半胱氨酸蛋白酶。 LAB胞外多糖还能抑制肿瘤血管生成。促进生长中的肿瘤血管化的主要因素是血管内皮生长因子(VEGF)。许多恶性肿瘤释放VEGF,以促进肿瘤内部及其附近微血管的形成。
肿瘤的血管化通过氧气和营养供应促进肿瘤的生长和发展。根据Deepak等人的研究,以5 mg/mL剂量给药L. acidophilus10307菌株的胞外多糖可导致促血管生成基因(包括VEGF基因)的显著下调,并刺激抗血管生成基因的表达。然而,该团队进行的另一项研究表明,相同的EPS也可以通过过表达EPO促进血管生成,EPO是一种刺激红细胞生成和提高癌细胞存活率的化合物。
鉴于这些不确定的结果,LAB胞外多糖在抑制肿瘤血管生成中的最终作用还有待解释。 众所周知,慢性炎症和癌变之间存在着很强的相关性。持续感染和慢性炎症,特别是在一些解剖部位(如肝脏和肠道),显著增加癌症发生的风险。LAB的各种代谢物(包括胞外多糖),被证明具有抗炎作用。
Bleau等人证明,鼠李糖乳杆菌RW-9595M菌株合成的EPS诱导的抗炎细胞因子(如IL-10)水平高于促炎细胞因子(如TNF-α和IL-6)水平。癌细胞被转录因子激活(主要是NF-κB),产生促炎细胞因子、趋化因子和生长因子。用EPS处理癌细胞系可通过抑制相应基因的表达而导致NF-κB的失活,同时增加IκBα抑制剂的浓度。
6.4 短链脂肪酸
LAB合成一系列对人体健康有益的代谢物。除了共轭亚油酸(CLA)和维生素B复合物外,短链脂肪酸(SCFAs)也被认为是LAB的重要代谢物。短链脂肪酸包括乙酸、丙酸和丁酸,这些化合物主要是在肠道细菌发酵膳食纤维过程中形成的。肠道SCFAs的总量很难量化,因为绝大多数SCFAs是由结肠细胞吸收和代谢的。
乙酸和丙酸主要运输到肌肉和肝脏,而丁酸则由结肠细胞原位代谢,占其能量需求的70%。Johansson等人的研究表明,健康志愿者以5×107 CFU/mL的剂量补充L. plantarum 299v持续21天,粪便中SCFAs的总浓度显著增加,并导致肠道内容物的pH值局部下降。
丁酸是LAB合成的最重要的短链脂肪酸。丁酸除了作为结肠细胞的主要能量来源外,还通过表观遗传学调控结肠黏膜正常细胞增殖和成熟相关基因的表达。丁酸还具有免疫调节、抗炎、抗氧化和抗癌活性。丁酸的抗炎和免疫刺激活性主要与阻断NF-κB的表达有关;NF-κB是一种转录因子,可控制许多炎症介质的表达,如细胞因子、酶、生长因子、热休克蛋白和免疫细胞受体。丁酸还能降低促炎细胞因子IFN-γ、TNF-α和TGF-β的表达,抑制一氧化氮合成的诱导,阻止淋巴细胞的增殖和活化。此外,丁酸还参与控制大肠中的氧化应激。 丁酸在预防结直肠癌的背景下被研究。
研究人员的兴趣与其对结肠细胞的双重作用有关;已知丁酸可调控正常结肠细胞增殖分化相关基因的表达,并抑制结肠癌细胞增殖,诱导早期凋亡。丁酸与结直肠癌细胞的GPR109A受体结合,诱导后者的凋亡。用丁酸盐孵育结直肠癌细胞可增加细胞内某些酶(超氧化物歧化酶2和过氧化氢酶)的表达,这些酶参与保护细胞免受活性氧的侵袭。
因此,丁酸对这些酶的刺激作用间接有助于抑制肿瘤生长。体外研究和动物实验都表明,丁酸会干扰癌细胞内的许多信号通路。 丁酸是由许多肠道细菌合成的,包括LAB。除丁酸外,双歧杆菌(Bifidobacterium spp.)发酵碳水化合物还会产生其他酸,如乙酸和乳酸,这些酸随后被其他肠道细菌(尤其是厚壁菌门细菌)转化为丁酸。
除了双歧杆菌外,其他重要的短链脂肪酸的生产者,如L. delubreckii subsp. bulgaricus和Streptococcus thermophilus产生丁酸和丙酸。García-Albiach等人证明,食用含有这些LAB菌株的酸奶对人体肠道产生丁酸盐具有促进作用。同样,Berni等人也发现补充益生菌鼠李糖乳杆菌GG(Lactobacillus rhamnosus GG)的儿童肠道微生物群中丁酸产生菌的比例更高。反之,根据Veiga等人的研究,摄入添加了Bifidobacterium animalis subsp. lactis的乳制品可促进大肠中其他合成丁酸盐的细菌数量的增加。
结论
乳酸菌(LAB)对人体健康的有益作用多年前就已为人所知;这些细菌提供抵抗病原体的保护,控制肠道上皮的完整性并刺激免疫系统。LAB是一种天然存在于发酵食品中的革兰氏阳性微生物,被用作益生菌,多年来一直受到研究人员的关注。LAB是具有多种功能和活性的生物活性化合物的有效来源,尽管尚未完全了解。
这些微生物合成的生物活性化合物包括短链脂肪酸、胞外多糖和细菌素等。最近的证据表明,LAB代谢物的活性不仅限于抗菌作用,而且还延伸到刺激宿主免疫和癌症控制。对LAB生物活性代谢物与免疫机制相互作用的研究表明,这些物质可以发挥强大的免疫调节作用,这也解释了其巨大的治疗潜力。LAB的抗癌活性在体外和动物模型中都得到了证实,可以对抗各种恶性肿瘤的癌细胞。
LAB通过多种机制抑制肿瘤生长,包括抗增殖活性、诱导细胞凋亡、细胞周期阻滞,以及抗诱变、抗血管生成和抗炎作用。越来越多的证据表明,LAB是一种很有前景的抗癌化合物来源,但这些发现需要在体外和动物模型中以及临床试验中进一步研究来验证。
不感兴趣
看过了
取消

人点赞

人收藏

打赏
不感兴趣
看过了
取消
©2012-2023 北京华媒康讯信息技术股份有限公司 All Rights Reserved. 注册地址:北京 联系电话:010-82736610
广播电视节目制作经营许可证 —(京)字第 17437号 京海食药监械经营备20200522号
京ICP备12011723号 京ICP证150092号
 京公网安备 11010802020745号
工商备案公示信息
互联网药品信息服务资格证书((京)-非经营性-2020-0015)
京公网安备 11010802020745号
工商备案公示信息
互联网药品信息服务资格证书((京)-非经营性-2020-0015)






您已认证成功,可享专属会员优惠,买1年送3个月!
开通会员,资料、课程、直播、报告等海量内容免费看!

打赏金额
认可我就打赏我~
1元 5元 10元 20元 50元 其它
打赏作者
认可我就打赏我~
扫描二维码
立即打赏给Ta吧!
温馨提示:仅支持微信支付!
已收到您的咨询诉求 我们会尽快联系您


































 010-82736610
010-82736610
 股票代码: 872612
股票代码: 872612




