
在高强度火灾后的第一个十年内,硝化作用的相对遗传能力相对较高,而自火灾以来,随着反硝化作用的相对遗传能力的增加,硝酸盐逐渐耗尽。
导读
野火在美国西部较为常见,会对植物的再生和简历产生有害影响,并严重影响碳(C)和氮(N)循环速率。土壤微生物在促进这些基本循环中起着关键作用,因此了解日益严重的火灾对土壤微生物群落的影响至关重要。本研究对原核生物(古细菌和细菌)的16S rRNA基因和真菌的ITS区域以及鸟枪法宏基因组学和微生物基因组重建进行高通量测序,并与先前发表的研究从内华达山脉中部火灾时间序列收集的土壤和植被数据配对,以考察高强度的火灾对土壤微生物群落的长期影响。
结果发现,由于地表植被、土壤化学成分和微生物群落组装过程的相关差异,高强度的野火会使得土壤微生物组通过数十年(>25年)才能恢复。我们的研究结果描述了火灾后土壤微生物中一个明显的分类和功能演替模式。
微生物组组成的变化与微生物功能潜力的变化相对应,特别是碳代谢的改变和氮循环潜力的增强,这分别与潜在的分解速率和无机氮可用性有关。基于宏基因组组装基因组(MAGs),本研究发现在最早的研究地点(自火灾发生4年以来)富集的细菌基因组具有独特的特征,如强烈的应激反应和降解热原多环芳烃C(pyC)的高潜力,使它们能够在火灾后的环境中茁壮成长。
本研究结果强调了土壤微生物群落在调节火灾后生态系统恢复中的重要性,并为将微生物生态学纳入火灾后森林管理提供了指导。
论文ID
原名:Ecological and genomic responses of soil microbiomes to high-severity wildfire: linking community assembly to functional potential
译名:土壤微生物组对高强度野火的生态和基因组响应特征:将群落装配与功能潜力联系起来
期刊:The ISME Journal
IF:10.302
发表时间:2022.4
通讯作者:Nicholas C. Dove
通讯作者单位:美国加州大学默塞德分校;美国橡树岭国家实验室
DOI:10.1038/s41396-022-01232-9
实验设计

图1. 内华达山脉中部火灾时间序列地图: A.符号表示采样点的大致位置,多边形为火灾的周界。每个采样点之间至少相距150 m,在空间尺 度上可能无法明显区分。采样点旁边的数字表示在该点的采样个数。B.每个时间序列采样点的平均植被覆盖类型百分比。覆盖类型有乔木(冷杉、黄松、栎树)、幼苗(黄松)、灌木(熊果属植物、柳树)、固氮植物(美洲茶属植物、蕨叶属植物)、草本植物(苔草属植物、禾本科植物)和裸土
结果和讨论
1. 高强度火灾导致土壤微生物群落多年恢复,并伴随地表植被和土壤化学成分的变化差异
微生物群落组成在火灾后各阶段均有显著差异(PerMANOVA-原核生物:p<0.001,R2=0.34;真菌:p<0.001,R2=0.18;图2,图S1,表S1)。即使在火灾后25年,微生物群落组成也与晚期演替森林之间存在显著差异(火灾后>115年,p<0.001)。
当把纬度和经度添加到该模型中时,发现只有经度是显著变化的(原核生物:p=0.008,真菌:p=0.009),但对微生物群落组成的影响可以忽略不计(R2均为0.02)。由于火灾的严重程度显著减缓了微生物的反应能力,因此数十年微生物群落组成的差异与火灾严重程度有关。
例如,火灾发生3年后,与受高强度火灾影响的土壤微生物群落相比,暴露于低强度火灾的土壤微生物群落的恢复能力更强。有研究表明,低强度的火灾根本不会影响到土壤微生物群落。历史上(前欧美殖民地)这一地区的火灾严重程度从低到重,每10到20年发生一次。
研究结果表明,高强度火灾后该生态系统的微生物群落的恢复时间比历史火灾恢复时间长,并且可能比历史微生物恢复时间更长。
图2. 内华达山脉火灾时间序列中微生物群落组成的控制:基于距离的冗余分析(dbRDA)对原核生物(A)和真菌(B)群落组成按土壤总碳(C)、总氮(N ) 、 硝酸盐 、 氨氮、磷酸盐(PO43−)、土壤pH值(1:2 w/v 0.01 M CaCl2)和持水量(WHC)。仅绘制了变化显著的土壤变量(p<0.05)。土壤测量方法是Dove等人使用相同的样本用于分子分析。点代表单个样本,并根据火灾时间和制备覆盖类型分别由不同颜色和形状表示。向量表示环境因子与dbRDA前两个轴相关性的方向和大小(由向量长度表示)。括号中的百分 比 为每个轴解释 的方差,需要注意不同的轴刻度。C.在每个时段内,整个群落原核生物的β-最临近分类指数(βNTI)的平均值(和标准误差,4-y : n=13、13-y:n=23、25-y:n =34,>115-y:n =20),它量化了观察到的零系统发育转换分布之间的偏差大小和方向,绘制为发生火灾以来时间的函数。βNTI的降低表明了更大的同质性选择。D.对于每个时间点内的原核生物(变量选择很难在图中的比例下查看),用系统发育箱方法计算的组装过程的相对优势。
不同植被覆盖类别(如乔木、幼苗、固氮植物、草本植物、裸土)的相对优势对火灾后微生物群落组成随时间变化的影响有限(图1B)。结果表明植物覆盖类型效应显著,但影响较小(PerMANOVA-原核生物:p=0.019,R2=0.06;真菌:p=0.016,R2=0.06,图2)。本研究没有发现火灾后时间和植被覆盖对微生物群落组成的影响之间存在显著的交互作用(原核生物:p=0.138;真菌:p =0.5 63)。
然而,树木/幼苗和草本/裸土植被组之间存在显著差异(表S2)。因此火灾后,随着时间的推移,树木的重建似乎部分与微生物恢复有关,不同主导真菌类型的转变可以证明。例如,在草本植被覆盖类型下,丛枝菌根真菌(AM)类群的相对丰度和 操作分类单位( OTU s)显著提高( ANOVA-相对丰度:p<0.001,丰富度:p=0.016,图S2A,B)。
此外,尽管外生菌根真菌(EM)类群的相对丰度在不同植被覆盖类型间差异不显著(p=0.256,图S2C),但EM的OTUs差异显著(p=0.017,图S2D),其中丰富度最高的是幼苗。EM分类群OTUs的相对丰度和丰富度自火灾以来随时间呈对数增加(相对丰度:p=0.003,rho=0.32,丰富度:p <0.001,rho=0.67),并在火灾发生13年后恢复到晚期演替时的相对丰度和丰富度(图S3)。
考虑到时间序列上的植被由与AM真菌共生的草本和固氮灌木(例如美洲茶属植物)转变为与EM真菌共生的乔木(如栎树和松树),因此优势菌根的变化可能反映了火灾后植被的演替。植物和微生物群落之间的关系也可能来自于在某些微生物的不同植被覆盖类型下发现的基质数量/质量和物理条件(如日照程度和土壤含水量)。事实上,火灾后早期演替植物的建立与微生物群落组成和功能相关。然而,我们在之前工作的基础上进一步表明,火灾后的地上和地下生物群的演替在干扰后的几十年内相互关联,特别是对于菌根真菌及其各自的宿主。
土壤化学成分随火灾时间的变化也可以解释微生物群落组成的变化(基于距离的冗余分析-原核生物:30%,p<0.001;真菌:18%,p<0.001;图2)。总体而言,土壤pH值(自火灾后随时间降低)对原核生物群落组成的预测力最强,土壤有机碳(SOC;自火灾后随时间增加)对真菌群落组成的预测力最强(表S3)。
微生物既控制着许多生物地球化学循环过程(如C和N循环),又对底物有效性的差异(如C和N循环)作出反应,因此很难判断微生物群落组成与土壤化学成分之间的相关性。然而,许多火灾后的生物地球化学变化是由非生物原因引起的,如SOC的燃烧、无机氮的释放和pH值的增加。在高强度的火灾下,土壤化学成分的变化可持续数十年,并影响微生物的长期演替。
2. 适应是关键:火灾发生后,选择随着时间的推移而增加
为了确定在这个时间序列中构建土壤原核生物群落的生态组装过程的影响,我们使用了两种零模型方法。这量化了确定性(即选择性)和随机组装对整个群落和相关分类的个体群落(即系统发育箱)的相对优势。第一种方法被用来扩展Ferrenberg和Knelman等人的建模结果,这些结果分别评估了野火在一年内和三年后对微生物群落组装的影响,并使用相同的零建模框架将其扩展到本研究的长期时间序列。
第二种方法使用与Stegen等人研究类似的框架,但已被进一步发展来计算不同组装过程对相关分类群的系统发育箱的优势度,这些优势度可能由不同施加于整个群落的因素决定的。我们将真菌排除在这项分析外,因为ITS基因区域在较大的系统发育距离中不一致,从而排除了使用基于ITS的系统发育树进行分析的必要性。
在第一种方法(即整个群落方法)中,我们发现决定性的同质选择, 一致性选择使微生物群落在环境条件下同质化,这是原核生物组装的主要驱动力(73-99%的范围,图S4A)。同质选择(βNTI<2)在恢复期间各时间点内均稳定增加(图2C)。先前使用相同建模框架的研究已表明,在野火发生4到16周后,基于小生境的选择性组装工艺最初较为宽松,但随后逐渐占据主导地位。
此外,在恢复3年后,89%的群落间的两两比较具有同质选择的特征。我们在之前的工作的基础上,展示了在与森林 恢复相关的较长时间尺度上,自火灾以来,选择的影响甚至随着时间的推移变得更加强烈( 图2C )。增加选 择很重要,因为这表明火灾后原核生物的组装和演替可以根据生态条件和微生物种群的遗传特征来进行预测。
环境 同质化的增加可能导致同质选择的主导。然而,不同与先前的研究,我们没有发现被烧毁地点的土壤化学具有更大的异质性。
例如,我们发现SOC和pH(解释微生物群落组成的两个最重要的土壤变量)的变异系数与火灾后的时间相似(表 S4 )。而pH值可能对微生物群落产生选择性压力,βNTI与土壤pH值的成对差异呈显著正相关(Mantel检验:p<0.001,rho=0.191;图S4B)βNTI与SOC的成对差异无正相关(Mantel检验:p=0.999;图 S4C )。
系统发育箱方法 在很大程度上能够支持整个群落方法的结果,随着时间的推移,每个时间点内的同质选择都在增加(图2D)。
然而,系统发育箱方法也检测到一个重要比例的随机装配因子,如 漂变 ,在4-y位点,自火灾以来随着时间的推移而减少(图2D)。在4-y位点,283个系统发育箱中 有136个的主要组装过程为漂变,其中以Delta-和γ-变形菌以及蓝细菌目吸血弧菌属为主。野火可能会减少微生物的丰度,从而增加“优先效应”的重要性。
事实上,在这些地点发生火灾后,微生物生物量C随着时间的推移而增加,表明野火减少了微生物生物量,并且这种影响是长期存在的。因此,高强度野火的急性干扰能力是一种重要的、慢性的选择力,在干扰数十年后还影响微生物群落的组成和组装过程。
3. 火灾后微生物C代谢的多年变化与潜在分解速率有关
在火灾发生后的30年内,微生物群落中C代谢所需的基因与晚期演替相比显著增加。 碳水化合物活性酶 (CAZy)基因的相对丰度自火灾以来,随着时间的推移而降低(p=0.011,rho=-0.618,图3A)。糖苷水解酶(GHs)的相对丰度在4-y位点达到最大值,并且从4-y到>115-y位点平均下降14%(p=0.009,rho= -0.631 ,图S5)。GHs基因相对丰度较高说明C可用性低,因为它代表了对 C获取的投入 。
例如,在根围隔实验中,相对于C含量丰富的根际土壤中发现的微生物基因组,在C含量相对贫乏的土壤中发现的微生物基因组富集了GHs。在另一个火灾后的森林中,火灾后3年增加了对C获取的投入。综上所述,这些结果表明,微生物对C获取的投入(可能以牺牲高效生长为代价)在火灾后相对较高,并在数十年内保持较高的水平(图3A)。
相对较大的 C获取的投入 可能是一种适应火灾的特性,因为在严重火灾下,容易降解的SOC会耗尽,导致C受限。然而,从CAZy丰度来阐明C的局限性应谨慎解释,未来的工作应将这些数据与转录组学和生理学测量相结合。
图3. 碳水化合物活性酶(CAZy)基因在野火时间序列中的分布和组成:A.由氨基酸(AA)编码读数校正的CAZy基因总丰度由箱线图表示,作火灾发生时间的相关函数。B.用 Bray-Curtis相异 数据归一化的主坐标分析(PCoA)表示CAZy基因组成的差异,括号中的百分比表示各轴解释的方差。C.箱线图,表示火灾以来的时间与功能分类的CAZy基因的归一化丰度相关的函数。橙色线表示显著(Spearman相关:p<0.05,n=4)关系的最佳拟合线性回归。注意不同的Y轴刻度。
微生物群落的底物选择性在火灾后的演替过程中也发生了变化,这表明CAZy基因的组成随火灾后的时间而变化(PerMANOVA:p<0.001,R2 =0.69,图 3B )。这表明,在火灾后生态系统恢复期间,微生物群落不仅在分类学上发生了变化,而且在新陈代谢上也发生变化。为了确定微生物C代谢如何在火灾后发生变化,我们还将根据C底物将一部分基因划分为功能组。C代谢的变化表现为自火灾以来,纤维素降解量随时间的增加而下降(p=0.018,rho=0.58,图 3C )。代谢能力的变化部分与底物及养分利用率的变化有关。
例如,我们还发现火灾后GHs的丰富度随着时间的推移而增加(p=0.030,rho=0.54),表明在火灾后恢复过程中C源的多样性增加。然而,火灾后纤维素的直接降解潜力更大,这可能不是纤维素底物增加的结果,因为它们通常在火灾后降低。
相反,较大的纤维素降解潜力可能是低碳、火灾后环境中碳获取的结果。纤维素具有比更复杂的C形式(如木质素)更大的底物利用效率,因此当C受限时,纤维素酶的产量应优于木质素分解酶。 为了了解CAZy基因如何影响生态系统过程,有必要将这些CAZy相对丰度与微生物群落的含量进行基准比较。因此,我们将本研究中的CAZy相对丰度乘以来自Dove等人研究中的相同样品的微生物量。我们发现自火灾以来,与CAZy基因相对丰度不同,这些归一化的CAZy相对丰度随时间的推移而增加(p =0.038,rho=0.52,图 4A )。
此外,我们还发现这些归一化的CAZy相对丰度与来自Dove等人的相同样品的28天实验室培养的CO2累积流量呈正相关(p < 0.001,rho=0.81,图 4B )。这表明,火灾后土壤中微生物生物量随时间的增加超过了早期演替微生物群落CAZy相对丰度的增加,从而影响了土壤的潜在分解速率。
然而,结果还表明,CAZy相对丰度以及微生物生物量的测量值可用于预测潜在的微生物分解。近期,研究人员对模拟火灾后的SOC再积累很感兴趣。我们的结果表明,这些模型可以通过结合微生物生物量和潜在的微生物遗传学来改进。
图4. 碳降解的遗传能力与二氧化碳(CO2)排放量有关:A.碳水化合物活性酶(CAZy)基因的总丰度说火灾后时间的相关性,通过氨基酸(AA)编码读数校正并乘以微生物量(MBC)与火灾后的时间。B.实验室培养28天的CO2累积排放与标准化CAZy相对丰度之间的相关性。橙色线表示显著(Spearman相关:p<0.05,n=4)关系的最佳拟合线性回归。
4. 土壤微生物氮循环能力的差异反映了火灾后土壤中无机氮的有效性
鉴于在许多陆地生态系统中,N是植物生产力的限制性营养元素,微生物N循环可能会影响生态系统的恢复。一般来说, N循环过程的相对遗传能力 (即关键N循环基因的相对丰度)在火灾后的前30年高于演替晚期(图5A)。这在生物固氮的相对遗传能力上表现得最为明显,火灾后4至25年比晚期演替相比平均高2.7倍(p =0.023)。这种遗传模式也被植物覆盖类型的测量结果所证实,表明共生固氮植物(如美洲茶属植物和蕨叶属植物)占这一演替时期植被覆盖的三分之二(图1B)。
然而,除了固氮之外,每个位点还具有特定的遗传N循环特征,对应于与火灾后生态系统演替相关的土壤和环境条件。例如,氨氧化的相对遗传能力在不同地点之间存在差异(p =0.016),并且在火灾后4年最高( 图5A )。此外,硝酸盐还原和一氧化二氮还原(反硝化的关键途径)的相对遗传能力在不同位点存在差异 (硝酸盐还原:p=0.043,一氧化二氮还原:p =0.015),并且在火灾后25年的地点最高(是晚期演替地点的两倍多,图5A)。
综上所述,这些结果表明,在高强度火灾后的第一个十年内,硝化作用的相对遗传能力相对较高,而自火灾以来,随着反硝化作用的相对遗传能力的增加,硝酸盐逐渐耗尽(图S6)。这些发现与来自多个生态系统的现场测量结果大体一致,表明硝化作用在火灾后的头几十年内得到了增强,从而为这些生物地球化学观测提供了遗传基础。
图5. 内华达山脉中部火灾时间序列的氮(N)循环潜力:A.对于每个路径,彩色圆圈代表相对于>115y位点的各位点之间的平均log2(倍数变化)。amo-氨单加氧酶(EC 1.14.99.39)、nap-硝酸盐还原酶(EC 1.7.1.2)、nar-硝酸盐还原酶(EC 1.7.99.4)、nif- 固氮酶 (EC 1.18.6.1)、nirB/D-亚硝酸盐还原酶大/小亚基(EC 1.7.1.15)、nirK/S、nor-亚硝酸盐还原酶前体(EC 1.7.2.1)、nos- 一氧化氮合酶 (EC 1.14.13.39)、nrf- 细胞色素c亚硝酸盐还原酶 (EC 1.7.2.2), nxr-亚硝酸盐氧化还原酶(EC 1.7.5.1)。B.火灾发生后N循环随时间变化的函数。线代表了无机N、硝化作用的相对遗传潜力和反硝化作用的相对遗传潜力随时间的相对差异。
主要N循环过程的相对遗传能力的演替变化与火灾后无机氮的有效性和时间有关。虽然这些不同N循环基因的相对遗传能力随火灾后的时间变化而不同,但所有测量的氮循环过程的生物量归一化遗传能力与火灾后的时间和无机氮的有效性性无关(p>0.05),该结果可能是由于这些功能基因的活性差异所造成的。当某些N循环基因由于有利的环境条件而相对富集时,它们的活性也会增加。因此,我们综合了无机氮有效性、其他环境因素(如C和O2)和N循环过程的相对遗传能力之间的关系,而不是生物量归一化的概念模型遗传能力(图5B)。
在干扰后的第一个十年内,N循环的特点是硝化程度相对较高,这可能是由于低植物N吸收率、低微生物固定铵(NH4+)和低C可用性,以 及由于 固氮灌木 丰 度在火灾后的增加使得N固定能力提高有关(图1B和5A)。此外,反硝化的遗传能力相对较低,并且硝酸盐(NO3-)在火灾后立即积累(图S6),其中低C可用性和O2可用性的潜在增加是限制环境因素。在连续的第二个十年中,硝化基因相对较少(图5A),这可能是由于植物对NH4+、C可用性和NH4+的微生物固定化的竞争加剧。这导致下游N循环过程的N相对较少,并且这些过程的相对遗传能力也降低(图5A)。
然而,随着火灾时间的推移,由于光照和湿度的限制,植物的生长和氮的吸收可能会减慢。这减少了对NH4+的竞争,有利于硝化作用和随后的反硝化作用,故这些过程的相对遗传能力增加(图5A)。因此,这种由由这些环境条件决定的相对较高的硝化和反硝化遗传能力的平衡保持了相对较低的NO3-水平(图S6)。
这些结果证明了土壤微生物群落、环境因素和循环过程之间的关系,并为先前发表的生物地球化学测量提供了急需的生物学基础,将微生物遗传学与生态系统功能联系起来。因此,火灾后微生物群落的演替在一定程度上决定了土壤的营养状况,并形成了火灾后生态系统恢复的轨迹。 5. 不同演替阶 段的微生物在分类学上是保守的,并适应火灾后的环境 我们得到了205个MAGs,约占宏基因组读数的20%,以评估 连续位点之间基因组解析的差异。本研究发现,自火灾发生以来,82个MAGs随时间推移大量减少,我们将其归类为早期演替,59个MAGs随着火灾后的时间而增加,将其归类为晚期演替(图6A;64个MAGs没有显示出任何明显趋势,表S5)。
图6. 早期和晚期演替宏基因组组装基因组(MAGs)的分类和估计生长速率:A.根据中心对数比(CLR)随火灾发生后的时间变化的丰度,将MAGs分为早期和晚期演替类群。B.火灾后平均 (±标准误差, n=4)MAGs增长率指数与时间的关系函数。C.各时间点的平均(±标准误差)生长速率指数分为早期和晚期演替(早期:n=82,晚期:n=59)。
早期和晚期演替MAGs的分组在分类学上是保守的(图6A)。该模式与我们的16S rRNA基因和鸟枪法宏基因组门谱一致,随着时间的推移,变形菌门和酸杆菌门增加,放线菌相对丰度降低(图S1),节杆菌属也是一样(图S7)。MAGs、鸟枪法宏基因组和扩增子数据集的分类学变化与先前关于各种生态系统野火后土壤微生物群落变化的研究一致,这表明至少在门水平上,野火后土壤微生物群落变化的许多情况是可推广的。
先前的研究表明,火灾后1到2年的微生物与其晚期演替对应的生物相比具有相对更快的增长率。然而,这些测量大多都是基于16S rRNA基因测序,它与最大生长速度相关,但不一定与实际生长速度相关。为了检验生长速率在本研究的演替梯度中是否随时间变化,使用 峰谷 比 (PTR)计算了演替位点上MAGs的原位生长速度。自火灾以来,个别MAGs估计的生长速度随火灾发生的时间而变化(MAGs同一性随机效应的混合模型:p =0.002)。
例如,13-y和25-y位点估计的生长速度大约是4-y位点的1.5倍(图6B)。然而,我们没有发现早期和晚期演替MAGs之间的平均估计增长率存在差异(Wilcox秩和检验:p =1.000,图6C)。这可能是由于火灾后生长速率随时间呈高斯分布,与火灾后10至30年相比,火灾后4年和>115年的增长率相对较低(图6B)。火灾后10到30年位点较高的增长率可能是由于土壤C(与4-y位点相比,表S4)和养分(与>115-y位点相比,图S6)相对较高的结果。传统的模式是,在演替早期(干扰后1到2年)微生物快速生长。然而,我们的研究结果表明,一旦微生物底物增加,演替中期微生物群落(干扰后10-30年)也具有相对较快的生长速度。这些结果并不一定会否定以前的模式,即早期演替微生物具有快速的增长率,因为火灾后4年(我们最早的火灾后测量时间)可能已 经太晚了,无法测量到 许多最 早的火灾后微生物。因此,为了了解火灾后早期微生物的演替,有必要在火灾后一年内进一步利用PTR和生理学方法(例如定量稳定同位素探测)研究微生 物的 生长速度 。
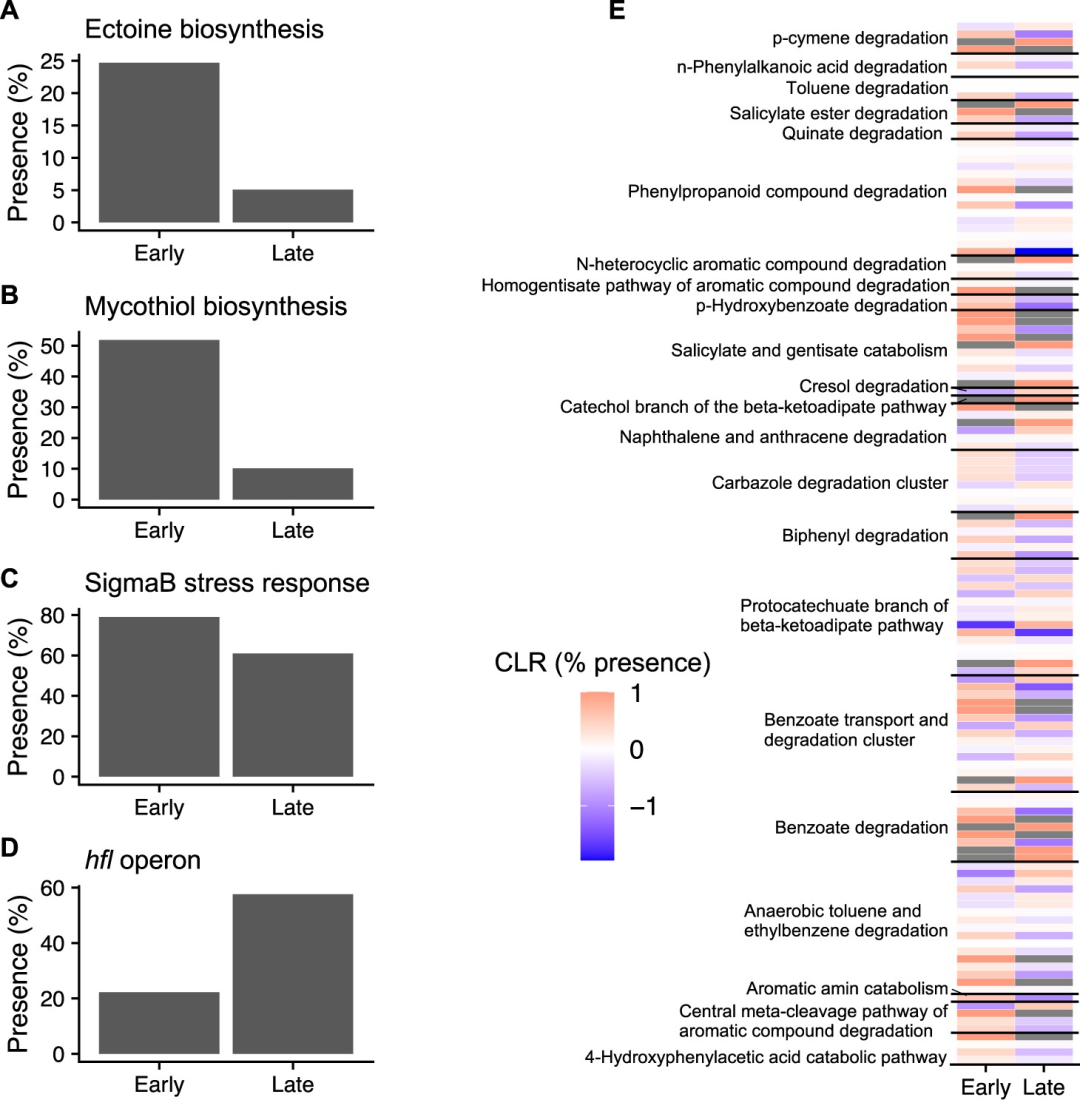
与晚期演替MAGs相比(图7A-C),早期演替MAGs富含应激反应基因(图7A-C),这使它们在火灾后的应激环境中生存下来。例如,早期演替MAGs具有 编码 四氢嘧啶 和放 线硫醇生物合成基因的可能性大约有五倍(卡方检验:p <0.05,图7A,B)。四氢嘧啶作为一种应激保护剂被释放,它具有渗透应激耐受性,对快速的温度波动提供保护作用,并保护DNA免受电离辐射损伤,均发生在火灾后树木树冠损失的环境中。 放线硫醇 是一种在放线菌中发现的谷胱甘肽类似物,可作为硫醇缓冲剂,维持细胞内的还原环境。
由于较低的持水能力(表S4)和沉积灰中发现丰富的金属氧化物,早期演替的位点可能具有高度氧化性。
因此,放线硫醇的产生可能是应对这种氧化还原应激的必要结果,特别是因为59个早期演替MAGs中有38个属于放线菌门,它们不合成谷胱甘肽。
此外,与晚期演替MAGs相比,与SigmaB应激反应相关基因在早期演替MAGs中也显著增多(p=0.033,图7C)。SigmaB是一种通用的、非特异性的应激调节因子,可激活多种基因以应对不同的应激反应,包括盐、热和渗透压力等。此外,晚期演替的M AGs富含hfl操纵子(p<0.001,图7D),其中包括hflX,一种使脓肿分枝杆菌对 大环内酯类抗生素 产生耐药性的抗性基因。
自火灾以来,微生物种群之间的竞争随时间的推移而增加,因此 可以针对抗菌素的细胞防御进行选择。综上所述,这些应激反应相关的基因在早期演替MAGs中富集,表明在高度氧化和干燥的火灾后的土壤中,提高细胞的适应能力是存活关键。
图7. 早期和晚期演替宏基因组组装基因组(MAGs)的特征。A-D.与压力相关基因的百分比含量。E .芳香碳降解主要途径中差异表达基因的热图(χ 2 检验- p <0.05,早期: n =82,晚期: n =59)。颜色代表每个基因百分比含量的中心对数比(CLR);灰色代表特定演替群体中不存在的基因(即CLR=未定义)。芳香碳降解主要途径中的基因见表S5。
早期 演替的MAGs还富含编码芳香族C降解的基因,这使得它们能够利用普通火灾后的C底物,并降解土壤中潜在的外源性化合物。多环芳烃C化合物是通过有机物的热解和不完全燃烧产生的,并且在火灾后的环境中大量存在。值得注意的是,这些MAGs通常具有芳香胺分解代谢、羟基苯甲酸酯降解和正苯基链烷酸降解的能力 (图7E,表S5) 。
这与近期研究结果相似,火丝菌属是一种常见的嗜热真菌,能够通过降解领苯二酚和奎宁酸来降解 pyC ,这一功能在我们早期演替的MAGs中也很普遍(图7E,表S5)。因此,本研究结果表明,早期演替的微生物特别适合降解pyC,潜在地提高了它们对没有这种能力的微生物的适应性。土壤中储存了54-109 Pg的pyC,微生物群落对pyC化合物的降解能力可能是决定火灾后土壤C库稳定性的关键因素。
由于本研究结果表明,火灾后微生物对其环境具有良好的适应性,因此,为了提高火灾后的管理目标,灾后微生物接种策略应优先考虑能够耐受火灾应激环境的微生物。虽然早期演替的微生物干预措施取得了好坏不一的结果,但本研究中发现火灾后微生物能够高度适应应激和氧化环境,这表明许多晚期演替的微生物可能无法在火灾后的早期演替环境中生存。因此,我们建议今后的微生物干预研究应侧重于促进培养或提取早期演替火灾后土壤的当地微生物种群,这些微生物种群具有强大的应激反应系统和合成应激保护剂的能力,如本研究中所强调的。
结论
这项研究表明,生态系统从高强度火灾中恢复可能需要几十年的时间,并与微生物群落的演替有关。具体来说,我们发现微生物遗传潜力的变化可以反映土壤氮的生物有效性,这可能会影响植物群落的建立和再生。
火灾4年后,微生物的恢复并不是由所有快速生长的原始物种决定的;相反,在生态系统演替早期和中期阶段典型应激、碳限制的火灾后土壤环境中,微生物组装主要由确定性选择决定的。
因此,今后的研究应该探讨火灾后土壤的管理,以及如何能够创造导致有益微生物(例如,固氮细菌、菌根真菌)恢复的条件,如本地植物群落的快速重建。在某些情况下,增加覆盖物可能有助于缓解碳限制和干旱胁迫环境对微生物群落的影响,提高微生物的适应能力和氮的稳定性。
然而,在大多数情况下,微生物接种剂的辅助定殖也许并不必要,因为分散限制在原核生物恢复中仅起次要作用,而菌根接种仍然可能有用;然而,目标真菌共生体应该具有强大的应激反应系统。随着全球范围内野火规模、严重程度和频率的增加,提高森林生态系统对火灾的抵御能力和制定有助于恢复的策略,对于灾后恢复和维持宝贵的森林生态系统服务至关重要。
不感兴趣
看过了
取消

人点赞

人收藏

打赏
不感兴趣
看过了
取消
©2012-2023 北京华媒康讯信息技术股份有限公司 All Rights Reserved. 注册地址:北京 联系电话:010-82736610
广播电视节目制作经营许可证 —(京)字第 17437号 京海食药监械经营备20200522号
京ICP备12011723号 京ICP证150092号
 京公网安备 11010802020745号
工商备案公示信息
互联网药品信息服务资格证书((京)-非经营性-2020-0015)
京公网安备 11010802020745号
工商备案公示信息
互联网药品信息服务资格证书((京)-非经营性-2020-0015)






您已认证成功,可享专属会员优惠,买1年送3个月!
开通会员,资料、课程、直播、报告等海量内容免费看!

打赏金额
认可我就打赏我~
1元 5元 10元 20元 50元 其它
打赏作者
认可我就打赏我~
扫描二维码
立即打赏给Ta吧!
温馨提示:仅支持微信支付!
已收到您的咨询诉求 我们会尽快联系您






































 010-82736610
010-82736610
 股票代码: 872612
股票代码: 872612




