
考虑到可能的治疗靶点和策略的数量巨大,以及患者本身和患者间免疫景观的差异,需要在在最能复制人类癌症的TME的小鼠模型,人多细胞类器官模型上进行仔细的临床前研究。
细胞因子及其受体作为肿瘤靶点或治疗策略已被广泛研究,如增强干扰素和白细胞介素(IL-2、IL-7等)的生长抑制、免疫刺激作用,或抑制TNF、IL-1β和IL-6等细胞因子的促炎、促肿瘤作用。新兴免疫疗法的出现,以及对肿瘤微环境(TME)的进一步了解,为利用细胞因子网络治疗癌症提供了新的方法,包括利用细胞因子增强其他疗法的活性或减轻免疫相关毒性。但许多挑战依然存在。2022年1月英国伦敦的研究人员于《Nature Reviews Cancer》杂志发表了一篇名为《Harnssing cytokines and chemokines for cancer therapy》的综述,作者对利用细胞因子及趋化因子治疗肿瘤的临床研究进行了系统性概述,现介绍如下:
细胞因子是由多种细胞分泌并参与细胞通讯的调节因子,主要包括白细胞介素、干扰素(IFN)、肿瘤坏死因子(TNF)超家族的一些成员、趋化因子和生长因子。白细胞介素在激活和调节先天性及适应性免疫反应中起着至关重要的作用。IFN除了具有强大的抗病毒作用,还具有一系列其他免疫刺激和抑制功能,以及抗肿瘤活性和自身免疫作用。TNF超家族由19种结构相关蛋白组成,具有促炎或抗炎活性。趋化因子由至少47种结构相关的细胞因子组成,这些细胞因子通常具有刺激细胞迁移的能力,与细胞外基质(ECM)相互作用从而控制细胞定向迁移。生长因子,在结构和功能上具有高度多样性,其表达通常由细胞因子诱导,某些生长因子如TGFβ家族成员,是有效的免疫抑制分子。
这篇综述主要强调细胞因子及其受体,而非相关的信号通路,特别关注结合细胞因子的联合疗法在实体肿瘤中的临床试验结果,同时该综述也讨论了从早期单药治疗的临床试验中吸取的经验教训。
TME中的细胞因子网络
细胞因子是TME中细胞通讯的关键介质。尽管IL-2、IFNα和IFNγ等细胞因子可促进TME中的抗肿瘤作用,但肿瘤细胞、免疫细胞和基质细胞导致的细胞因子失调可以出现在肿瘤发生的所有阶段以及治疗过程当中。因此,利用细胞因子的免疫刺激效应和纠正细胞因子失调有利于开发癌症治疗的潜力。某些细胞因子与肿瘤的发生、进展和转移密切相关(图1框1),促炎因子的异常产生是细胞癌变的常见结果。估计有10%–25%的癌症源于慢性炎症性疾病,但癌症也会引起TME的炎症改变,尤其是促肿瘤细胞因子的产生。
多项研究表明,人类癌症的TME中细胞因子及其受体非常丰富。一项荟萃分析对TCGA数据库中超过10000个肿瘤(包括33种不同的癌症类型)进行了分析,揭示了6种不同的免疫亚型:伤口愈合型、IFN-γ优势型、炎症型、淋巴细胞耗竭型、免疫平静型和TGF-β优势型。免疫亚型主要由细胞因子、趋化因子及其受体的不同mRNA表达谱确定。重要的是,基于细胞因子和趋化因子信号网络的肿瘤亚型分类可能会影响细胞因子相关疗法的选择。例如,TGFβ受体拮抗剂可能适用于TGFβ优势亚型,而某些白介素或白介素-刺激细胞因子可能适用于淋巴细胞耗竭亚型。
总之,迄今为止研究的所有TME中细胞因子及其调节途径的复杂网络在众多实体肿瘤中是共享的,尽管这些网络组成可能因个体免疫环境不同而存在差异。鉴于细胞因子在免疫反应中的作用以及在恶性肿瘤中的失调表现,我们在此介绍利用细胞因子或其拮抗剂用于癌症治疗的方法。
图1 细胞因子在肿瘤微环境中的作用
框1 细胞因子的潜在促肿瘤作用
细胞因子疗法的临床试验
强有力的证据表明,一些细胞因子尤其是与适应性免疫应答有关的细胞因子(如IFNγ、IL-2和IL-12),有助于抗肿瘤免疫。在过去的40年里,人们尝试利用这些细胞因子治疗癌症(表1)。
IL-2是调节先天性和适应性免疫的关键细胞因子,也是T细胞增殖、存活和分化的关键介质,是CD4 +T细胞和NK细胞的生长因子,并参与B细胞产生抗体。IL-2的免疫刺激特性为30多年前开始的临床试验提供了理论基础。高剂量重组IL-2已分别于1992年和1998年被FDA批准用于治疗转移性肾细胞癌(RCC)和黑色素瘤。约12%的黑色素瘤患者和约7%的RCC患者达到完全反应(CRs)。临床上应用IL-2的缺点包括半衰期短(约85min),需要每8小时给药一次,以及严重的并发症如血管渗漏综合征和心脏毒性等。此外,IL-2可诱导Treg细胞活性从而消除抗肿瘤反应。IL-2对Treg细胞的影响以及血管渗漏综合征的高风险尤其与其受体的高亲和力三聚体(IL-2Rαβγ)有关。相比之下,CD8+ T细胞、记忆T细胞和NK细胞表达缺乏α亚单位(CD25)的低亲和力二聚体IL-2受体(IL-2Rβγ)。人们已经利用这种差异研发出可以选择性结合低亲和力IL-2Rβγ的药物。例如,bempeg是IL-2的聚乙二醇化形式,优先结合IL-2Rβγ,并通过β亚单位(CD122)激活CD8+ T细胞和NK细胞,而非Treg细胞。这种药物的聚乙二醇化也可能减少与外周血T细胞活化相关的毒性,并增加IL-2的半衰期(约20小时)。另一种基于IL-2R的治疗方法包括使用抗CD25单克隆抗体(mAb)去除免疫抑制性Treg细胞。
表1 肿瘤治疗策略中的主要细胞因子
IL-7是白介素共同γ链受体家族的成员,其通过IL-7Rα和γc的异二聚体受体传递信号。IL-7具有多种免疫效应,包括在不诱导Treg细胞的情况下诱导幼稚和记忆性T细胞增殖。临床试验中,尽管观察到循环CD4+和CD8+ T细胞水平以及T细胞受体(TCR)多样性增加,但IL-7在肿瘤患者中没有疗效。临床前模型中,重组人IL-7或IL-7与IgG Fc片段的融合形式(以延长其半衰期)可潜在地诱发炎性TME。因此,多个试验正在联合免疫检查点抑制剂(ICIs)对此类药物进行研究,但结果尚未报告。
IL-15在临床前模型中,IL-15可以促进NK细胞和CD8+ T细胞的生成、增殖和活性。已有170多个有关IL-15的临床试验在进行中,结果显示IL-15单药治疗疗效有限。IL-15半衰期短,但与高亲和力受体IL-15Rα结合时更稳定。因此,由IL-15与截短形式的IL-15Rα复合物组成的IL-15超抗原已经进入临床研究阶段,并取得了良好的初步结果(表1)。进行最广泛试验的超抗原N-803已经与其他免疫疗法联用进行试验。在80例非肌层浸润性膀胱癌(NMIBC)患者中,72%的患者在膀胱内加入N-803后产生完全反应(CRs),且多数患者有持久性反应(中位持续时间为19.2个月)。
IL-12能刺激NK细胞和CD8 +T细胞增殖,促进Th1细胞的分化和IFNγ的产生从而促进抗原提呈。此外,IL-12还具有强大的抗血管生成作用,因此是一种重要的潜在治疗性细胞因子。与IL-2类似,IL-12的短半衰期和剂量毒性限制了临床应用(框2)。事实上,针对各种重组IL-12的试验,无论是作为单一疗法,还是与其他药物联合使用,都已证明对患有皮肤T细胞淋巴瘤、艾滋病相关卡波西肉瘤、非霍奇金淋巴瘤或RCC的少部分患者有反应,但严重的不良事件如死亡等导致了该疗法的弃用。
为了避免毒性,NHS-IL12通过将两个IL-12分子融合到针对DNA-组蛋白复合体的抗体中,这使得细胞因子能够选择性地输送到肿瘤坏死区域。此外,细胞因子也可与抗体片段相连,而非整个mAbs。如IL12-L19L19由IL-12和来自mAbL19的串联Fv片段组成,而mAbL19针对在肿瘤中高表达而在正常组织中不表达的纤维连接蛋白外域B剪接变异体。IL12-L19L19目前正在晚期实体肿瘤或弥漫性大B细胞淋巴瘤患者中进行临床试验(NCT04471987)。肿瘤内注射编码IL-12的质粒可导致IL-12的局部持续表达,并使黑色素瘤患者的注射部位和远处的肿瘤消退。目前多项试验正在评估上述质粒运输系统,包括腺病毒载体和嵌合抗原受体(CAR)T细胞。
IL-10通常被认为是TME中的免疫抑制因子,但IL-10通过增加CD8 +T细胞活化,在各种肿瘤模型中显示出抗癌作用。研究者使用聚乙二醇化IL-10制剂对晚期实体肿瘤患者进行了一系列试验。据报道,pegilodecakin单药治疗可增加活化的浸润性CD8 +T细胞的数量。在多队列I期IVY研究中,在20例重度预处理的RCC患者中,有4例在pegilodecakin单药治疗后观察到部分反应(PRs),但未诱发细胞因子释放综合征(CRS),尽管中位无进展生成时间(PFS)仅为1.2个月。对于标准治疗无效的其他实体瘤患者,pegilodecakin单药治疗可使小部分患者的病情持久稳定。尽管转移性胰腺导管腺癌(PDAC)患者有良好的反应,但在二线FOLFOX化疗中添加pegilodecakin并不能改善最终III期试验的客观缓解率(ORR)、PFS或总生存率(OS)。
白介素作为过继细胞疗法的辅剂 采用过继性细胞疗法如淋巴细胞浸润疗法治疗黑色素瘤,需要细胞因子支持输入细胞的存活和增殖。IL-2在这种情况下通常作为辅助药物使用,同样,IL-2也经常用于CAR-T疗法。CAR-T细胞除了能表达针对肿瘤抗原的特异性受体外,还能分泌细胞因子。事实上,分泌IL-12或IL-15的CAR-T细胞已在晚期实体肿瘤患者中进行临床试验(如NCT03721068和NCT02498912),这些细胞可能在不诱导Treg细胞的情况下促进TME中T细胞的细胞毒性和持久性。临床前研究中,表达IL-7和CCL19的CAR-T细胞增强了抗肿瘤活性。因此,在淋巴瘤和实体瘤中的试验正在评估双重表达IL-7和CCL19的CAR-T细胞疗法的效果(NCT03198546、NCT04381741和NCT03929107)。
IFNα包括13种密切相关的抗病毒细胞因子。干扰素的许多活性支持其在癌症治疗中的应用(图1):上调MHC分子,促进DC细胞迁移,增强细胞毒性T细胞活性,抗血管生成,并可直接对肿瘤细胞产生毒性或抑制作用。此外,其他有效的癌症治疗措施,包括化疗、免疫治疗和靶向治疗等,通常依赖于干扰素信号,尤其是cGAS- STING信号通路。
尽管IFNα在循环中的半衰期很短,但IFNα疗法的系统性不良事件(包括疲劳和抑郁)发生率高(框2)。因此IFNα单一疗法很少用于治疗实体肿瘤,该疗法目前已被新的免疫疗法和靶向治疗取代。然而,这种方法在治疗血液系统恶性肿瘤方面确实具有一定的临床活性和实用性。
框2 基于细胞因子治疗策略的失败原因
IFNγ是先天性和适应性免疫系统的主要调节因子,由NK细胞、NKT细胞、CD8+T细胞和CD4+T细胞产生,巨噬细胞和B细胞是其作用的主要靶细胞和效应细胞。研究已证实IFNγ通过CD4+T细胞和CD8+ T细胞产生抗肿瘤作用。然而,IFNγ尚未被FDA批准用于治疗任何实体肿瘤,相关研究只显示了轻微的临床疗效。
在卵巢癌患者中,腹腔注射IFNγ与显著的肿瘤消退有关,皮下注射IFNγ在一线化疗方案的基础上改善了PFS。据报道,IFNγ联合卡铂-紫杉醇的一线化疗方案可以缩短OS。在最近的一项试验中,用聚乙二醇化IFNα-2b和IFNγ体外治疗自体单核细胞,然后将其腹腔注射给铂类难治性卵巢癌患者,目的是通过干扰素使TME偏向M1/Th1表型, 11例可评估患者中有2例出现PR。在NMIBBC中,膀胱内注射IFNγ可诱导肿瘤消退,但效果不如BCG标准疗法。IFNγ在其他实体肿瘤患者中的试验,包括转移性RCC,结肠癌,黑色素瘤,小细胞肺癌和晚期PDAC,均显示无效。
值得注意的是,在一些实体肿瘤中,IFNγ信号的基因表达特征与对ICIs的反应性呈正相关。此外,对抗CTLA4单抗无反应的黑色素瘤患者,通常存在IFNγ通路的缺陷。研究还表明,IFNγ可诱导肿瘤MHC表达和T细胞浸润。所有这些发现都支持重组IFNγ与ICIs联用进行临床试验 (NCT02614456和NCT03063632)。
细胞因子受体拮抗剂的临床试验
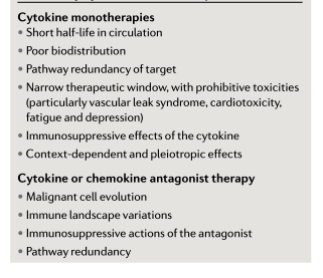
随着细胞因子阻断疗法的广泛应用,特别是针对TNF、IL-6和IL-1的疗法在一些炎症性疾病和自身免疫性疾病中的应用,临床前研究为细胞因子阻断剂用于癌症治疗提供了理论基础。特别是在2000-2010年,大量研究表明,针对关键炎性因子的单抗或受体拮抗剂在异种移植或基因工程模型中具有抗癌活性。此外,炎性细胞因子及其信号通路缺乏小鼠的肿瘤进展和转移的风险降低。总而言之,这些发现表明,靶向TME中的细胞因子网络可以重塑炎症细胞浸润状态或阻止炎症细胞迁移到肿瘤部位。临床前数据还表明,某些细胞因子拮抗剂可以重塑TME为使其成为抑制肿瘤的微环境,从而促进肿瘤特异性适应性免疫反应并抑制转移扩散。
这些发现促进了临床试验的展开。大多数抗细胞因子单抗的临床试验都是针对晚期癌症患者的早期剂量安全性研究。作为单一疗法,这些药物尽管耐受性很好但疗效有限或无效。因此,这些拮抗剂目前正与其他免疫疗法相结合(表2),以下讨论了一些可能令人感兴趣的研究。
表2 肿瘤治疗策略中的主要细胞因子及其受体阻断剂
TNF拮抗剂 TNF是一种由单核细胞、巨噬细胞以及包括成纤维细胞和内皮细胞在内的其他细胞在应激反应中产生的一种促炎因子。TNF具有多种促肿瘤活性,如刺激细胞因子级联反应、促进纤维化和改变黏附受体,在化学致癌的早期阶段起重要作用。抗TNF抗体或可溶性TNF受体融合蛋白是几种常见的治疗自身免疫性或炎症性疾病的有效方法 (表2)。在一项涉及晚期癌症患者的I期临床试验中,抗TNF单抗英夫利昔单抗耐受性良好,没有剂量限制的毒性。给予英夫利昔单抗后1h后,血清TNF中和,血浆CCL2、IL-6和血清CRP水平下降。41例患者中有7例患者病情稳定(持续10-50周以上),没有任何患者有疾病加速进展的证据。在对37例耐药或细胞因子治疗(TNFα和/或IL-2)无效的肾癌患者进行的两项序贯II期研究中,应用标准剂量或大剂量的英夫利昔单抗时有3例患者出现PR,14例患者病情稳定。临床前研究表明,抗TNF药物可增强ICIs的活性和/或降低自身免疫性不良事件。
IL-6拮抗剂 IL-6-JAK-STAT3信号通路具有重要的免疫调节作用。IL-6在TME中具有促炎和促肿瘤作用 (图1)。针对IL-6及其受体或JAK-STAT3通路的药物已被FDA批准用于治疗炎症或骨髓增生性肿瘤以及由CAR-T诱导的CRS(表2)。有三种IL-6拮抗剂被临床批准使用:siltuximab(抗IL-6 mAb)、tocilizumab和sarilumab(抗IL-6R mAbs)。临床前研究表明,中和IL-6或抑制JAK-STAT3下游信号具有强大的抗癌活性,尽管这些效果尚未转化为肿瘤患者的临床获益。Castleman病是一种罕见的由IL-6驱动的良性淋巴增生性疾病,临床试验显示抗IL-6或抗IL-6R单抗对该肿瘤和临床症状有持久反应。然而,其他包括多发性骨髓瘤、前列腺癌、肾癌和卵巢癌在内的临床试验,只显示了单药疗法的轻度生物学活性。IL-6拮抗剂如tocilizumab通常用于缓解一些接受CAR-T疗法并发生严重CRS的患者。T细胞刺激的巨噬细胞和单核细胞是产生IL-6的主要细胞,尤其是在肿瘤负荷较高的患者中,强烈的CAR-T细胞激活可能导致主要由IL-6参与的大规模潜在致命性细胞因子风暴。
IL-1β拮抗剂 与TNF和IL-6类似,大量临床前研究强调了IL-1α和IL-1β的促肿瘤作用。IL-1β主要由TME中的髓系细胞和成纤维细胞产生,尤其是通过炎症小体刺激其他炎症介质、细胞侵袭和免疫抑制(图1)。Anakinra是一种天然存在的IL-1R拮抗剂的重组形式,它竞争IL-1α和IL-1β与IL-1R1的结合,从而抑制生物活性。该药通常用于治疗痛风和类风湿性关节炎,以及因IL-1信号失调引起的罕见炎症性疾病。Anakinra常被加入到现有的治疗方法并用于包含结肠癌、胰腺癌和乳腺癌在内的实体肿瘤患者的临床试验中(表2)。Anakinra联合5-FU和贝伐单抗治疗转移性结直肠癌的II期试验显示,中位PFS和OS持续时间分别为5.4和14.5个月。在一项涉及11名转移性HER2−乳腺癌患者的初期试验中,Anakinra导致白细胞炎症转录信号的持久变化:IL-1和NF-κB信号下调,T细胞和NK细胞毒性相关基因上调。此外,研究表明抑制IL-1可能会减轻CAR -T疗法的神经毒性。Anakinra治疗严重的CAR -T相关毒性的大规模队列研究正在进行中(例如,NCT04432506、NCT04359784、NCT04148430和NCT04205838)。
TGF-β抑制剂 TGF-β在TME中具有免疫抑制和诱导纤维化的作用(图1),临床前研究表明,当TGF-β拮抗剂与ICIs联合使用时,抗肿瘤活性增强。其主要作用机制似乎是通过改变TME中ECM来增强T细胞浸润。TGF-β的临床靶标主要是利用TGF-βRI丝氨酸/苏氨酸激酶域的小分子抑制剂,如Galunisertib,或抗TGF-β单抗,如fsolimumab (表2)。其他靶向方法包括针对TGF-β受体和TGF-β配体。Bintrfusp-α是一种抗PD-L1-TGF-β RII融合蛋白,能够同时靶向PD-L1和所有TGF-β亚型,但目前有关bintrafusp的各项试验已经终止。将TGF-β拮抗剂与ICIs相结合的许多其他研究尚未进行报道。
趋化因子受体拮抗剂的临床试验
刺激免疫细胞定向迁移是趋化因子及其受体家族中最突出的作用。癌症中趋化因子网络存在失调,趋化因子可以促进新生血管的生成,它们的趋化活性经常由肿瘤细胞所决定(图1)。因此,临床前研究支持利用一些趋化因子及其受体作为治疗靶点在实体肿瘤中开展临床试验进行进一步探索(表3)。
表3肿瘤治疗策略中的主要趋化因子及其受体阻断剂
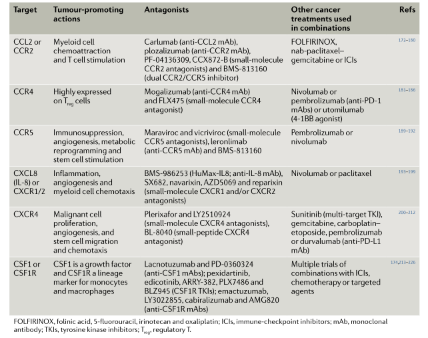
抑制CCL2-CCR2轴 趋化因子受体CCR2及其配体CCL2是髓系细胞向多种肿瘤趋化的重要介质;在临床前模型中,阻断CCL2-CCR2轴可抑制肿瘤相关巨噬细胞(TAM)的募集。针对CCR2或CCL2的各种药物已经进行临床试验,但大多数试验结果显示无效,多种药物已被放弃,包括卡鲁单抗、多氮卓单抗和PF-04136309。
第二代CCR2抑制剂CCX872-B与FOLFIRINOX应用在一项涉及晚期PDAC患者的Ib期试验中, 18个月的OS为29%,结果优于单独使用FOLFIRINOX。此外,小分子双重CCR2/5拮抗剂BMS-813160目前正处于I/II期测试,既可以作为单一疗法也可以与化疗药物或nivolumab联用,用于晚期微卫星稳定(MSS)的结直肠癌或晚期肺癌(NCT03184870)以及局部晚期或边缘可切除的PDAC(NCT03496662和NCT03767582)。
CCR4拮抗剂 CCR4是由Treg细胞表达的CCL17和CCL22的受体。多种CCR4拮抗剂已经纳入临床试验中(表3),迄今为止在肿瘤学中唯一被批准的CCR4拮抗剂是用于治疗T细胞淋巴瘤的莫伽珠单抗。FLX475是一种耐受良好的口服小分子CCR4拮抗剂,具有良好的药代动力学特性。该药无论是作为单一疗法,还是与培溴利珠单抗(NCT03674567)联合使用,目前均在进行I/II期试验。尚未发表的数据表明,单一疗法和联合疗法都有药物反应
CCR5拮抗剂 CCR5有多个配体,包括CCL3、CCL4、CCL5和CCL8。CCR5信号在促进肿瘤生长方面具有多样性:免疫抑制、刺激血管生成、代谢重编程和干细胞扩增(图1)。目前至少有四种CCR5拮抗剂正在进行临床试验:BMS-813160、小分子抗逆转录病毒药物maraviroc和vicriviroc,以及抗CCR5单抗leronlimab (表3)。这些试验目前还未显示有效结果。
IL-8和CXCR1/2抑制剂 CXCR1/2是刺激血管生成、炎症、抑制髓系细胞趋化和肿瘤干细胞存活的关键受体(图1)。IL-8(CXCL8)是CXCR1/2的主要配体之一,这种促炎因子在许多实体肿瘤中过表达,并具有多种促肿瘤活性。此外,血清IL-8浓度增加与肿瘤负荷和对ICIs的耐药性相关。研究表明,阻断IL-8-IL-8R轴可增强ICIs的疗效。抗IL-8单抗BMS-986253正在多项涉及实体肿瘤的早期试验中进行评估,主要与nivolumab联合使用(例如NCT03689699)。此外,针对IL-8R的药物,包括CXCR1/2双重抑制剂SX682、NAVARIXIN以及CXCR2拮抗剂AZD5069,正在与各种抗PD-1或抗PD-L1单抗 (表3)一起进行评估。另一种CXCR1/2的口服抑制剂Reparixin与紫杉醇联合用于转移性HER2−乳腺癌或TNBC的患者,尽管没有观察到主要药物毒性,但没有可信的证据表明药物活性增强。
CXCR4拮抗剂 CXCR4是一种趋化因子受体,在超过75%的癌症中都有过度表达,在肿瘤的增殖、扩散和血管生成中发挥重要作用。多种实验小鼠模型中CXCR4拮抗剂都能抑制肿瘤生长。plerixafor是临床中应用最为广泛的CXCR4拮抗剂,该药能动员造血干细胞进入外周血因而被用于非霍奇金淋巴瘤或多发性骨髓瘤。许多其他CXCR4拮抗剂已经进入临床开发阶段 (表3)。在类似的情况下,CXCR4配体CXCL12或CXCL7的拮抗剂正在临床试验评估中。无论如何,尚无可信的证据表明CXCR4拮抗剂可以增强ICIs临床反应的预期效果。
CSF1R和CSF1拮抗剂 CSF1R是髓系细胞的谱系标志,也是生长因子CSF1(M-CSF)的受体。Pexidartinib是一种口服酪氨酸激酶抑制剂,其活性主要是针对CSF1R,一项III期试验的数据显示其在肌腱滑膜巨细胞瘤患者中有良好反应,目前已获得FDA的批准。此外,多种抗CSF1R和抗CSF1单抗的临床试验正在进行中,包括LY3022855、AMG820和PD-0360324等,无论是作为单药治疗还是与化疗、ICIs或其他靶向药物联合使用。
总结
自从细胞因子疗法首次应用于癌症患者以来已经有40年了。在随后的几年中,我们对细胞因子家族、抗肿瘤免疫、TME和癌症演变的了解大大增加。我们现在可以得出结论,抑制或增强单个细胞因子或趋化因子途径不太可能对晚期癌症具有持续的活性,只有少数患者除外。事实上,除了黑色素瘤和肾癌的IL-2和IFNα的显著例外,基于单剂细胞因子的治疗在临床上并不有效,尽管IL-2的一些改良的受体选择性形式和新型趋化因子导向的拮抗剂在这方面具有潜力。临床疗效的缺乏部分与大多数细胞因子治疗的局限性有关,包括短半衰期、狭窄的治疗窗口和免疫抑制机制的上调(框2)。因此,这些治疗已被ICIs和其他新型免疫疗法所取代。细胞因子治疗的当前临床研究方向集中于与其他免疫疗法的结合和/或细胞因子的修饰,以改变其药代动力学和结合亲和力,从而降低毒性并避免免疫抑制因子(如Treg细胞)的增加;上述IL-2“超因子”以及IL-12免疫细胞因子最能说明后一种策略。
利用过去40年中获得的所有知识,短期使用细胞因子或其抑制剂暂时改变TME中免疫细胞的活性或ECM的产生可能是最有希望的,无论是与其他疗法结合还是在(新)佐剂环境中。在考虑细胞因子或细胞因子拮抗剂的联合和序贯治疗时,值得注意的是,某些化疗可降低全身炎症细胞因子水平,而高循环水平的IL-8与ICIs的耐药性相关。相反,卵巢癌患者的(新)辅助化疗激活了TME中的IL-1β–NLRP3炎性体途径,有免疫原性细胞死亡的证据,表明在癌症治疗的某些阶段使用特定细胞因子拮抗剂时要谨慎。考虑到可能的治疗靶点和策略的数量巨大,以及患者本身和患者间免疫景观的差异,需要在在最能复制人类癌症的TME的小鼠模型,人多细胞类器官模型上进行仔细的临床前研究。
编译:王沛,翁梅琳
审校:张军,缪长虹
参考文献:Propper, D. J., & Balkwill, F. R. (2022). Harnessing cytokines and chemokines for cancer therapy. Nature reviews. Clinical oncology, 19(4), 237–253.
不感兴趣
看过了
取消

人点赞

人收藏

打赏
不感兴趣
看过了
取消
©2012-2023 北京华媒康讯信息技术股份有限公司 All Rights Reserved. 注册地址:北京 联系电话:010-82736610
广播电视节目制作经营许可证 —(京)字第 17437号 京海食药监械经营备20200522号
京ICP备12011723号 京ICP证150092号
 京公网安备 11010802020745号
工商备案公示信息
互联网药品信息服务资格证书((京)-非经营性-2020-0015)
京公网安备 11010802020745号
工商备案公示信息
互联网药品信息服务资格证书((京)-非经营性-2020-0015)






您已认证成功,可享专属会员优惠,买1年送3个月!
开通会员,资料、课程、直播、报告等海量内容免费看!

打赏金额
认可我就打赏我~
1元 5元 10元 20元 50元 其它
打赏作者
认可我就打赏我~
扫描二维码
立即打赏给Ta吧!
温馨提示:仅支持微信支付!
已收到您的咨询诉求 我们会尽快联系您





































 010-82736610
010-82736610
 股票代码: 872612
股票代码: 872612




