
Pi 和 Ca 的代谢不能相互隔离。它们在 ECF、肠道、骨骼和肾脏的运输机制中密切相互作用。
Phosphate Metabolism in Health and Disease
Peacock M. Phosphate Metabolism in Health and Disease. Calcif Tissue Int. 2021 Jan;108(1):3-15. doi: 10.1007/s00223-020-00686-3. Epub 2020 Apr 7. PMID: 32266417.
健康和疾病中的磷酸盐代谢
磷是一种原子量为 31 的 5A 元素,占植物和动物重量的 0.6% 以上。三种同位素可用于研究磷代谢和动力学。31 P 是稳定的,而放射性同位素33 P 的半衰期为 25 天和32P 的半衰期为 14 天。磷酸酯和磷酸酐是常见的化学键,磷是有机分子中的关键元素,参与多种基本细胞功能。这些包括通过三磷酸腺苷 (ATP) 进行的生化能量转移、通过核苷酸 DNA 和 RNA 维持遗传信息、通过环磷酸腺苷 (cAMP) 进行细胞内信号传导以及通过甘油磷脂实现膜结构完整性。然而,本综述侧重于作为弱酸的无机磷 (Pi) 的代谢。磷酸具有与氧相连的所有三个氢,是一种弱二质子酸。它有 3 个 pKa 值:pH 2.2、pH 7.2 和 pH 12.7。在生理 pH 值为 7.4 时,Pi 以 H 2 PO 4 (-)和 HPO的形式存在4 (2-)并充当细胞外液 (ECF) 缓冲液。Pi是跨组织隔室和细胞运输的形式。生物体液中 Pi 的测量基于其与不测量有机磷的钼酸铵的反应。在人类中,体内 80% 的磷以磷酸钙晶体(磷灰石)的形式存在,它赋予骨骼和牙齿硬度,并作为主要的磷储存器(图 1 ) )。其余部分存在于软组织和 ECF 中。膳食磷,包括无机和有机形式,在上消化道中被消化。吸收的 Pi 以由 ECF Pi 浓度、血流速率和细胞 Pi 转运蛋白活性决定的速率进出骨骼、骨骼肌和软组织以及肾脏(图 2 )。在生长过程中,磷净增加,随着老化,磷净损失发生。骨磷储层被总磷需求耗尽和补充。骨骼肌富含磷,用于基本的生化能量转移。肾脏是 ECF Pi 浓度的主要调节器,因为它具有处于密切内分泌控制下的肾小管对 Pi (TmPi) 的最大重吸收能力。它也是通过尿液排出的 Pi 过剩的主要排泄途径。跨细胞和细胞旁的 Pi 转运是通过广泛分布于组织中的多种转运机制进行的,在肠道、骨骼和肾脏中尤为重要。Pi 转运蛋白受激素轴的调节,该激素轴包括成纤维细胞生长因子 23 (FGF23)、甲状旁腺激素 (PTH) 和 1,25 二羟基维生素 D (1, 25D)。Pi 和钙 (Ca) 代谢密切相关,临床上两者都不能孤立地考虑。Pi 代谢疾病影响骨骼,如骨软化症/佝偻病,软组织如异位矿化,骨骼肌如肌病,肾脏如肾钙质沉着和尿路结石形成。
成人中磷的含量:骨骼、软组织和细胞外液(克,对数刻度)。显示钙的相应数据以供比较
磷酸盐 (Pi) 以 mg/24 小时进出组织隔室。在 1400 毫克的膳食磷中,1120 毫克在上肠被 ECF 吸收,210 毫克通过内源性分泌返回肠道,导致 910 毫克净 Pi 吸收和 490 毫克粪便排泄。在骨骼中,180 mg 通过骨形成沉积,180 mg 通过骨吸收返回 ECF。在肾脏,5040 mg 在肾小球被过滤,4130 mg 通过肾小管重吸收返回 ECF,其中 910 mg 从尿中排出。在软组织中,Pi 在 ECF 和细胞之间交换
介绍
磷是一种原子量为 31 的 5A 元素,占植物和动物重量的 0.6% 以上。三种同位素可用于研究磷代谢和动力学。31 P 是稳定的,而放射性同位素33 P 的半衰期为 25 天和32P 的半衰期为 14 天。磷酸酯和磷酸酐是常见的化学键,磷是有机分子中的关键元素,参与多种基本细胞功能。这些包括通过三磷酸腺苷 (ATP) 进行的生化能量转移、通过核苷酸 DNA 和 RNA 维持遗传信息、通过环磷酸腺苷 (cAMP) 进行细胞内信号传导以及通过甘油磷脂实现膜结构完整性。然而,本综述侧重于作为弱酸的无机磷 (Pi) 的代谢。磷酸具有与氧相连的所有三个氢,是一种弱二质子酸。它有 3 个 pKa 值:pH 2.2、pH 7.2 和 pH 12.7。在生理 pH 值为 7.4 时,Pi 以 H 2 PO 4 (-)和 HPO的形式存在4 (2-)并充当细胞外液 (ECF) 缓冲液。Pi是跨组织隔室和细胞运输的形式。生物体液中 Pi 的测量基于其与不测量有机磷的钼酸铵的反应。在人类中,体内 80% 的磷以磷酸钙晶体(磷灰石)的形式存在,它赋予骨骼和牙齿硬度,并作为主要的磷储存器(图 1 ) )。其余部分存在于软组织和 ECF 中。膳食磷,包括无机和有机形式,在上消化道中被消化。吸收的 Pi 以由 ECF Pi 浓度、血流速率和细胞 Pi 转运蛋白活性决定的速率进出骨骼、骨骼肌和软组织以及肾脏(图 2 )。在生长过程中,磷净增加,随着老化,磷净损失发生。骨磷储层被总磷需求耗尽和补充。骨骼肌富含磷,用于基本的生化能量转移。肾脏是 ECF Pi 浓度的主要调节器,因为它具有处于密切内分泌控制下的肾小管对 Pi (TmPi) 的最大重吸收能力。它也是通过尿液排出的 Pi 过剩的主要排泄途径。跨细胞和细胞旁的 Pi 转运是通过广泛分布于组织中的多种转运机制进行的,在肠道、骨骼和肾脏中尤为重要。Pi 转运蛋白受激素轴的调节,该激素轴包括成纤维细胞生长因子 23 (FGF23)、甲状旁腺激素 (PTH) 和 1,25 二羟基维生素 D (1, 25D)。Pi 和钙 (Ca) 代谢密切相关,临床上两者都不能孤立地考虑。Pi 代谢疾病影响骨骼,如骨软化症/佝偻病,软组织如异位矿化,骨骼肌如肌病,肾脏如肾钙质沉着和尿路结石形成。
循环和 ECF 中的磷
循环和 ECF 负责 Pi 进出参与 Pi 代谢的器官的运输(图 2 )。在血浆中,磷含量约为 12 mg/dL (3.87 mmol/L),其中约三分之一以无机磷 (Pi) 的形式存在(图 3 )。超过 80% 的血浆 Pi 是非蛋白质结合的,包括三种质子化物质以及与 Ca、镁和钠的各种复合物 [ 1 ]。尽管血浆 Pi 有大约 20% 的蛋白质结合,但由于 Donnan 膜效应,超过 95% 是可超滤的。因此,通常通过测量血浆中的总 Pi 浓度来评估 Pi 转运到肾小球滤液中的情况。只有在酸碱显着变化的情况下,这个假设才成立。血浆和其他生物体液中的 Pi 通过与钼酸铵反应形成磷钼酸盐来测量,其在 340 nm 处的吸光度与 Pi 的浓度成正比。成人血浆 Pi 范围为 2.5 至 4.5 mg/dl [0.81–1.45 mmol/L]。在儿童中,由于 TmPi 较高,该值较高,并且随着年龄的增长,从新生儿的约 6 mg/100 mLGF 逐渐降低到成人参考范围 [ 2 ](图 4 )。血浆 Pi 浓度具有可遗传成分 [ 3 , 4 ],随 PI 摄入量增加 [ 5 ],并具有昼夜变化 [ 5 , 6 ](图 5 )。成人参考范围男女相同,不受年龄影响。循环中的 Pi 浓度是评估 Pi 代谢的重要指标。低磷血症和高磷血症总是反映潜在的疾病。急性低磷血症和高磷血症是重症监护病房的常见发现,导致重症患者的高发病率 [ 7 , 8 ](表 1 )。慢性低磷血症会导致肌肉骨骼疾病(表 1 )。它导致儿童骨矿化延迟产生佝偻病和成人骨软化症,并导致近端肌肉无力,这反映了 Pi 在骨骼肌中化学能转移中的关键作用。相反,慢性高磷血症表现为皮下、血管和神经组织中的软组织矿化(表 1 )。肾功能衰竭是慢性高磷血症的最常见原因,并且具有复杂的病理生理学 [ 9 ]。一个潜在的异常是磷酸钙使 ECF 过饱和,血浆总 Pi x Ca 浓度乘积已被用于估计这种风险。根据八钙离子产物的评估,血浆对于磷酸钙的沉淀是亚稳态的。在大约 10 mg/dL 的正常总血浆 Ca 浓度下,在 Pi 的血浆浓度低于 14 mg/dL,即乘积约为 140 时,预计不会发生磷酸钙的自发沉淀。事实上,在患有70 mg 2 /dL 2的乘积是预期值的一半,清楚地表明还涉及晶体形成的各种组织促进剂。然而,该产品并不能比单独的血浆 Pi 浓度更好地预测 [ 10 ]。
图 3
人血浆中的磷含量。总磷约为 12 mg/dL,其中 72% 为有机磷,28% 为无机磷;无机磷 [磷酸盐] 包括 20% 的蛋白质结合和 80% 游离,包括各种离子种类。从 (1) 中重新绘制的数据
图 4
在 291 名女孩和 273 名男孩中,禁食过夜后肾小管最大重吸收磷酸盐 [TmPi] mg/100 ml 肾小球滤过液 [GF] 与年龄的关系为 6 -18 岁。从 (2) 中重新绘制的数据
图 5
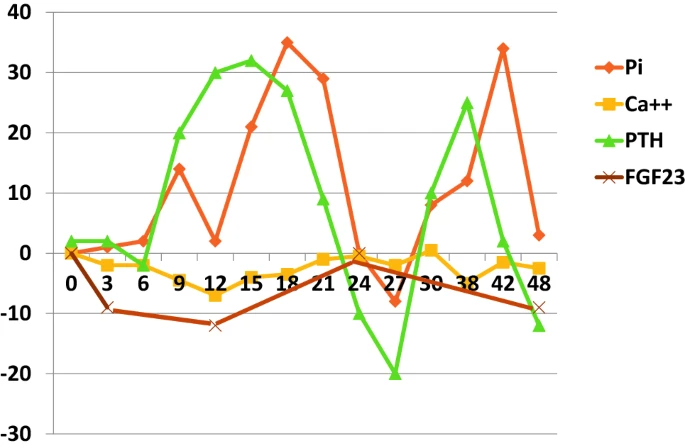
8 名健康男性 48 小时内血清磷酸盐 (Pi)、离子钙 [Ca++]、甲状旁腺激素 [PTH] 和成纤维细胞生长因子 23 [FGF23] 的昼夜变化,相对于基线的百分比变化。血清 Pi 的范围为 40%,而 Ca++ 为 8%;FGF23 的范围为 10%,而 PTH 为 50%。数据重绘自 (6)
表 1 与 a:慢性低磷血症,b:急性低磷血症,c:高磷血症相关的条件和机制
膳食磷
饮食是体内磷的主要来源。从饮食历史和营养数据库评估磷营养。出于营养需求和代谢平衡研究的目的,有机和无机磷的膳食摄入量通常基于 24 小时昼/夜期间消耗的食物。磷存在于大多数食物中,并且与蛋白质含量非常相似,在美国,每克蛋白质摄入约 15 毫克磷 [ 11 ]。大约 30% 的膳食磷以动物肉和蔬菜的形式食用。另外 30% 作为乳制品消费,其中约 70% 是无机磷。其余 40% 以 Pi 的形式摄入食品、食品加工的食品添加剂和膳食补充剂 [ 11 ](图 1)。 6 )。在食品标签、口服补充剂和药物中,磷含量通常不存在或记录不充分。与化学分析相比,使用营养数据库对膳食磷含量的严重低估进一步加剧了这种不确定性 [ 12 ]。每天 700 毫克的推荐每日摄入量 (RDA) 和每天 580 毫克的估计平均需求量 (EAR) [ 13 , 14 ] 在美国饮食中通常超过大约两倍 (表 2 ) [ 15 ]。EAR 有点武断,因为慢性膳食磷不足导致骨软化症/佝偻病不容易评估。每天 4000 毫克的可耐受膳食上限 (UL) 也值得怀疑,因为有证据表明,在某些情况下,如此高的摄入量可能会促进骨骼和心血管疾病 [ 16 ]。磷以 Pi 的形式被吸收。然而,磷的吸收生物利用度并不容易评估,因为它取决于消化的完整性、饮食中各种 Pi 结合剂的存在以及食物来源。例如,磷含量高的膳食植酸盐(肌醇六磷酸)的摄入量约为每天 1 克,素食饮食中的摄入量可能要高得多 [ 17 ]。植酸盐中的磷可通过植酸酶的消化被植物和反刍动物利用。然而,在人体中,生物利用度是可变的,因为烹饪会破坏植物植酸酶,人体消化酶缺乏植酸酶,并且植酸在人体小肠的 pH 值下大部分处于固态,其中大部分 Pi 被吸收。此外,一些口服药物是强 Pi 结合剂。例如,因胃酸而长期摄入的氢氧化铝会与饮食中的 Pi 结合,导致 Pi 耗竭并导致骨软化症 [ 18 ]。事实上,在慢性肾功能衰竭中,口服 Pi 结合剂在治疗上被摄入以降低 Pi 的生物利用度并预防高磷血症 [ 10 , 19 ]。
图 6
膳食磷食物来源 (11)
表 2 男性/男孩和女性/女孩的每日磷摄入量和 NHANES 2005-2006 的 RDA
肠道:Pi 吸收
小肠具有很大的 Pi 运输能力,负责吸收大部分膳食磷。吸收效率约为 80%,其中约 20% 的吸收的 Pi 作为内源性分泌物从 ECF 返回到肠腔(图 2 )。吸收通过细胞旁扩散和活跃的、可饱和的跨细胞机制发生,这些机制在低磷摄入时起作用 [ 20 , 21 ]。由于饮食中的磷通常处于过剩需求中,因此扩散在人类中占主导地位。然而,所有涉及的机制及其相对大小尚未完全阐明。众所周知,Pi 从肠腔转运到肠细胞是通过两个钠依赖性溶质载体家族发生的(图 7 )。II 型转运蛋白 NaPi2b [SLC34A2] 主动跨细胞转运 Pi [ 22 ]。它存在于肠细胞的顶膜中,在低腔内 Pi 浓度下起作用,其活性受 1,25D/VDR 调节。然而,该途径在数量上的重要性仍然不确定,因为人类 SLC34A2 的失活突变不会导致磷酸盐代谢的重大障碍,尽管它们确实会导致肺泡磷酸钙微石症 [ 23 ]。PiT1 [SLC20A1] 和 PiT 2 [SLC20A2] 也参与了肠上皮细胞对 Pi 的细胞内转运 [ 24 ]。另一方面,细胞旁 Pi 转运受肠上皮细胞中质子浓度的调节,该浓度由 SCLC9A3 编码的钠氢交换器 NHE3 调节。对 Tenapanor(一种设计用于在肠道局部起作用的小分子)的研究表明,细胞旁途径是一种主要的肠道 Pi 转运机制,可以通过药理学进行操作 [ 25 ]。通过抑制 NHE3 的活性,Tenapanor 减少钠转运到肠上皮细胞中,并增加质子浓度。细胞 pH 值的增加选择性地增加了对 Pi 转运的细胞间紧密连接阻力并减少了 Pi 肠道吸收(图 7 )[ 26 ]。在肾功能衰竭和 Pi 潴留的患者中,Tenapanor 治疗可显着降低血清 Pi [ 27 ]。Pi 吸收可以通过放射性标记吸收测试在人体中测量 [ 28 ]。它在许多疾病中减少,包括慢性肾功能衰竭、小肠吸收不良、维生素 D 缺乏性骨软化症、低磷性骨软化症,并在原发性甲状旁腺功能亢进和特发性肾结石形成中增加(图 8 )[ 20 ]。在这些疾病中,Pi 吸收分别反映了低和高 1,25D 血清浓度,强调了 125D/VDR 在 Pi 吸收中的核心作用。小鼠肠道中的 VDR 敲除使 Pi 吸收效率降低了 50% [ 29 , 30 ]。然而,低膳食 Pi 摄入量对增加吸收的影响 [ 31 , 32 ] 似乎与 1,25D/VDR 轴无关 [ 29 ]。FGF23 对人体 Pi 吸收的影响主要通过对 1,25D 分泌的主要影响而间接影响 [ 33 ]。Pi 的内源性分泌可以通过平衡和动力学研究进行评估,并且很大程度上取决于 ECF Pi 浓度(图 2 )。粪便中出现的未吸收磷是通过化学方法测量的,但除了代谢性磷平衡[ 12 ]外,几乎没有提供有用的临床信息。
图 7
膳食消化液中的磷酸盐 [Pi] 通过小肠中的跨细胞和细胞旁转运机制吸收。NaPi2b 和 PiT1 转运蛋白调节肠细胞中的跨细胞转运。1,25D/VDR 促进 Pi 传输。NHE3 转运蛋白通过设置控制细胞旁通道活性的细胞内质子浓度来调节细胞旁 Pi 转运。FGF23/Klotho-FGFR 在 Pi 吸收中的作用(如果有的话)是未知的
图 8
正常受试者 [n = 受试者人数] 和慢性肾功能衰竭、小肠吸收不良、维生素 D 缺乏性骨软化症、低磷血症性骨软化症、原发性甲状旁腺功能亢进症患者的放射性磷吸收 [ 50 mg Pi 中的32 P,分数速率/小时] (PHPT) 和特发性肾钙结石病。改编自 (20)
骨骼:Pi 运输
骨骼是最大的 Pi 储存库,重要的是也是调节 Pi 转运的主要激素 FGF 23 的来源。骨矿物质 Pi 可与 ECF Pi 交换,但这种通量对ECF Pi 稳态没有贡献 [ 34、35 ] 。骨骼中的 Pi 有两种基本的运输途径。第一个涉及 ECF Pi 进出骨骼的运输。第二个是骨内 Pi 从成骨细胞转运到磷灰石晶体。Pi 从 ECF 到新骨形成并通过骨吸收返回 ECF 的每日净转运量少于肾脏和肠道(图 2 ) )。骨转换在骨和 ECF 之间转移 Pi,并控制新骨的形成和不需要或受损骨的吸收。它可以促进骨骼生长、骨折修复和适应机械需求。由 sclerostin 途径调节的骨形成和由 RANKL 途径调节的骨吸收通常是紧密耦合的 [ 36 , 37 ]。大约 180 mg Pi 沉积在地层中,每 24 小时通过再吸收去除 180 mg。启动重塑周期的骨骼的去除由破骨细胞执行。它吸收类骨质并通过溶解磷灰石将 Pi 释放回 ECF 池。在骨形成和吸收部分分离的状态下,例如生长或骨骼固定,骨和 ECF 之间的 Pi 分别净增加或减少。已经提出了除骨转换以外的机制,允许骨库将 Pi 转移到 ECF 和从 ECF 转移。在需要大量 Pi 和 Ca 在相对较长的时期内同时排出骨库的状态下,例如怀孕和哺乳, 38、39 ] 。_ 一旦怀孕和哺乳完成,这个过程就会逆转,耗尽的矿物质会被完全替代。然而,并非所有研究都证实发生了这种变化 [ 40 ],溶骨性骨溶解的机制仍有待阐明。ECF Pi 浓度和 Pi 在骨骼中的内部转运对于磷灰石的形成都是必不可少的。ECF 中的 Pi 浓度部分决定了 Pi 进出骨细胞的速率,骨细胞是负责维持骨骼健康的细胞、成骨细胞、负责骨形成的细胞和负责包裹骨组织的内层细胞。在严重的慢性低磷血症中,例如肿瘤引起的骨软化症 [ 41 ]、XLH [ 42 ] 和缺乏维生素 D 的骨软化症 [ 43 ],从 ECF 到矿化前沿的 Pi 降低到矿化速度严重减慢的程度,导致儿童佝偻病和成人骨软化症(表 1 )。在骨骼内,Pi 也需要由成骨细胞生成和运输以形成磷灰石。磷灰石 [Ca 10 (PO 4 ) 6 (OH) 2 ]的成熟晶体,大约 3 nm 厚,大约 50 x 25 nm 的长度和宽度,被分层成胶原蛋白支架 [ 44 ]。它们还与类骨质蛋白和骨细胞及其小管在解剖学上密切接触(图 9 )。 PiT1&2 [ 45 , 46 , 47 , 48 ]调节进出骨细胞的活跃 Pi 转运。在成骨细胞中,Pi 被运输到包含形成无定形 Ca-Pi 所需的机制的基质囊泡中 [ 49 , 50 ](图 10 )。基质囊泡含有孤儿磷酸酶 1 (PHOSPH1),一种对 Pi 产生至关重要的酶 [ 51 ]。大小约为 200 nm 的微泡被胞吐到细胞外基质中,新生的无定形 Ca-Pi 晶体在其中吸积更多矿物质,形成成熟的磷灰石晶体。类骨质基质流体中的 Ca-Pi 过饱和,不需要的矿化被许多抑制剂控制。碱性磷酸酶和外核苷酸焦磷酸酶磷酸二酯酶 [ENPP1] 调节焦磷酸盐(一种关键抑制剂)在类骨质基质中的浓度,同时调节 Pi 转运至基质囊泡的可用性 [ 52 , 53 , 54 ]。在没有 PHOSPH1 或碱性磷酸酶活性的情况下,骨骼不能以正常速率矿化,导致佝偻病和骨软化症,而在没有 ENPP1 的情况下,会发生婴儿期广泛的动脉钙化。 由金属内肽酶 PEX [ 52 ]调节的同胞蛋白也是关键的晶体抑制剂,作用于多个位点以防止在不需要的位置形成晶体,例如骨细胞小管 [ 55 , 56 ]。因此,ECF 的 Pi 供应不足或成骨细胞 Pi 转运失败都会导致矿化缺陷,导致骨软化症和佝偻病。
图 9
磷酸盐 [Pi] 进入骨骼并在骨骼内。Pi 通过 Pit1 和 PiT2 从 ECF 转运至成骨细胞。在成骨细胞中,Pi 与钙 [Ca] 一起被包装在微泡 [MV] 中并被胞吐到骨基质中。新生的磷灰石晶体与新形成的胶原纤维对齐。进一步向成熟磷灰石晶体中的矿物质增加受磷酸盐/焦磷酸盐 [Pi/PPi] 比例和抑制蛋白(包括牙本质基质蛋白 1[DMP] 和骨桥蛋白 [OPN])的调节。新形成的基质与骨细胞及其小管网络密切相关
全尺寸图片
图 10
基质囊泡磷酸盐[Pi]来自三个来源。在囊泡内,孤儿磷酸酶 1[PHOSPH1] 从磷酸胆碱 [PC] 中释放 Pi。在囊泡外,由组织非特异性碱性磷酸酶 [TNAP] 和三磷酸腺苷 [ATP] 上的外核苷酸焦磷酸酶磷酸二酯酶 [NPP1] 的活性释放的 Pi 通过 PiT1 转运到囊泡中。后者增加骨基质中的焦磷酸盐 [PPi] 浓度
肌肉:Pi 运输
自主肌肉不参与 Pi 的整体运输和代谢(图 2 )。然而,将其包括在此处是因为近端肌病是慢性低磷血症的一个突出特征,而横纹肌溶解会导致急性高磷血症。通过双 X 射线吸收测定法 (DXA) 测量的男性瘦体重约为 20 kg/m 2,女性约为 15 kg/m 2 [ 57 ]。由于体积大,随意肌含有大部分软组织磷。它主要以有机磷的形式存在,特别是ATP和磷酸肌酐。肌肉中的细胞内游离 Pi 约为 1-2 mg/dL [3-5 mmol] [ 58 ],与 ECF Pi 呈线性相关 [ 59 ] ]。Pi 转运到肌肉细胞受 Pit1 和 PIT2 转运蛋白的调节 [ 60 ]。正常的 ECF 和肌肉细胞 Pi 浓度对于维持磷酸肌酐的储存和作为肌肉机械活动能量来源的 ATP 的功能至关重要 [ 61 , 62 ]。由于缺乏维生素 D 的骨软化症和肿瘤诱导的骨软化症引起的肌病对治疗反应迅速 [ 43 ],这表明肌肉中的 Pi 供应受损是主要的病因。
肾脏:Pi 重吸收和尿液排泄
肾脏处理日常 Pi 运输的主要部分(图 2 ),也是 1,25D(图 11 )的来源,它是调节 Pi 运输的关键激素。肾小球滤过每 24 小时将超过 5,000 mg Pi 输送到近端小管,近端小管将其中的 80% 以上重新吸收回 ECF(图 2 )。肾脏是控制循环中 Pi 浓度的主要器官 [ 63 ]。它通过肾小球滤过率 (GFR) 和近端小管对 Pi 的重吸收率来执行此功能。由于 TmPi 较高,儿童的 Pi 浓度高于成人。随着 GFR 的降低,与慢性肾功能衰竭一样,Pi 向近端小管的输送速率也相应降低。除非 Pi 肠道吸收补偿性减少,和/或 Pi 肾小管重吸收减少,和/或净组织 Pi 增加增加,否则循环中的 Pi 浓度不可避免地升高。由于饮食中的磷通常过量,肠道无法补偿 GFR 的慢性下降。除生长期间外,软组织和骨骼中 Pi 的增加是有限的。因此,GFR 下降的补偿很大程度上取决于肾小管减少重吸收的能力。 60、64、65、66、67 ] ( 图 11 ) 。 _ _ _ _ _ 使用 NaPi 2a 特异性抑制剂的研究表明,NaPi2a 起主要作用 [ 68 ]。Pi 通过包括 XPR1 在内的转运蛋白从小管细胞分泌回 ECF [ 69 ]。近端小管是 Pi 重吸收的主要部位。它的管状最大 Pi 重吸收能力 (TmPi) 约为 2 mg/dL,超过此值的过滤后 Pi 的任何增加都会从尿液中排出 [ 70 ]。有一个范围约为 1 mg/100 mL GF 的张开,在此范围内肾小管重吸收逐渐增加,然后达到完全 TmPi。临床上最好通过测量血浆 Pi 和血浆肌酐 (Cr) 以及相应的尿 Pi 和尿 Cr 在不超过 1 至 2 小时的适当时间间隔内收集血液中点的血液来计算 Pi 的最大肾小管重吸收的尿液收集。不应使用 24 小时的时间间隔,因为血液 Pi 浓度在 24 小时内变化很大,导致不正确的低 TmPi。禁食过夜后收集血液和尿液,以减少饮食和 GFR 的可变影响并优化个体的可重复性。肾小管重吸收可表示为与肌酐清除率相关的 Pi 排泄分数(血浆 Pi × 尿 Cr/血浆 Cr × 尿 Pi)。由于 GFR 在表达 TmPi 的个体内部和个体之间存在差异,因此每单位肾小球滤液 (TmPi/GFR) 提供的 TmPi 估计值可以更准确地在个体内部和个体之间进行比较。基于来自健康受试者磷酸盐输注的数据,TmPi 可以从列线图导出 [ 71 ] 或来自等式,例如 TmPi = P - P E /1–0.1 log e (P/P E ) 其中 Pi = 血浆 Pi,P E = 尿 Pi x 血浆 Cr/尿 Cr [ 63 ]。TmPi 在新生儿中较高,并随着年龄的增长逐渐降低,直到青少年达到成人值(图 4 )。尿液 Pi 可以通过多种方式表达。作为 Pi/Cr 比率,它是 TmPi 计算的一部分。对于营养研究、代谢平衡研究和尿液结石形成风险的估计,使用 24 小时内收集的尿液中的 Pi。在磷平衡的成年人中,24 小时尿 Pi 提供了膳食磷摄入量的估计值 [ 72 , 73 ]。 在 Pi 正平衡 [ 12 ]的成长儿童中,或在摄入 Pi 粘合剂 [ 19 ] 的受试者中,24 小时尿 Pi 几乎可以减少到零,情况并非如此。24 小时尿 Pi 仅与膳食钙摄入量呈弱正相关。然而,当摄入大量钙补充剂时,例如,在患有骨质疏松症或甲状旁腺功能减退症的患者中,钙作为磷结合剂,24 小时尿磷会降低 [ 74 ]。在儿童骨骼成熟之前,24 小时尿 Pi 没有性别差异。此后,由于较高的膳食 Pi 和 GFR,男性的 24 小时尿 Pi 绝对值高于女性,尽管他们的尿 Pi/Cr 比率相同。24 小时尿 Pi 被测量为评估肾结石形成的泌尿风险因素的浓度 [ 75 ]。在健康受试者中,Ca-Pi 结晶尿很常见,可以顺利通过,这反映了尿液 pH 值对 Ca-Pi 过饱和的主要影响。因此,虽然 Pi 是泌尿系结石的常见成分,但它的存在依赖于 pH 值,并不反映 Pi 代谢异常。磷酸钙结石在原发性甲状旁腺功能亢进症患者中占主导地位,在特发性结石病患者中以草酸钙/磷酸盐混合结石为主,在尿液感染患者中以磷酸镁铵结石为主 [ 76 ]。重要的是,Ca-Pi 作为集合管中的晶体聚集体或作为肾乳头中的上皮下沉积物(兰德尔斑块)作为肾结石形成的病灶起着关键作用 [ 77 ]。
图 11
约 80% 的肾小球滤过磷酸 [Pi] 通过近端小管中的跨细胞机制被重吸收。NaPi2a 和 NaPi2c 调节 Pi 转运进入小管细胞,小管细胞转运通过细胞并通过 XPR1 转运到 ECF。近端小管具有 PTHR1 和 FGFR/Klotho 受体。远端小管具有 FGFR/Klotho 和酶 CYP27B1 和 CYP24A1 的受体,负责将 25 羟基维生素 D 代谢为 1、25 二羟基维生素 D 和 24、25 二羟基维生素 D
Pi的内分泌调节:FGF 23、PTH、1,25D。
Pi 代谢的内分泌调节是通过三种激素 FGF 23、PTH 和 1,25D 的高度整合作用进行的 [ 78 , 79 ]。 骨细胞 [ 81 , 82 ]分泌的 FGF 23 [ 80 ]发挥着核心作用(图 12 )。ECF Pi 浓度在几天内增加 [ 72 , 73 ] 但不是在几小时内急剧增加 [ 83 ] 上调 FGF 23 分泌,进而通过近端小管中的 FGFR/Klotho 受体 [ 84 , 85 ] 降低 NaP2a 和 NaP2c 的活性。从管腔液到 ECF 的跨细胞 Pi 转运减少,导致 TmPi 降低和 ECF Pi 降低 [ 86 ]。同时,肾脏 1,25D 产量下降导致 Pi 肠道吸收减少 [ 80 , 87 ]。总体效果是预防高磷血症并维持正常磷血症。相反,Pi ECF 的减少会下调 FGF 23 分泌,导致 TmPi 增加和 Pi 吸收增加。总体效果是预防低磷血症并维持正常磷血症(图 12 )。高磷摄入导致的 FGF23 增加比摄入减少导致的增加更显着,这表明 FGF23 的主要功能是防止高磷血症和异位矿化 [ 72 ]。骨细胞感知 ECF Pi 浓度的机制仍未确定 [ 88 , 89 ]。它不涉及快速作用的正反馈循环 [ 82 ],而是涉及 ECF Pi [ 72、73 ]的慢性(数天)变化。血清 Pi 的昼夜变化与血清 FGF23 的增加无关,而血清 Ca 的昼夜变化与血清 PTH 的变化成反比 [ 6 ](图 5 )。这种相对较慢的正反馈回路响应可能在于 Pi 传感机制。骨细胞敲除研究有证据表明,被高浓度 Pi 磷酸化的 FGFR1 可能是 Pi 传感机制的一部分 [ 90 ]。此外,PiT 2 [ 91 ] 和 AMP 激活激酶可能参与传感机制 [ 92 ]。非激素刺激的 FGF23 分泌在低铁状态下发生(图 12 )[ 93、94、95 ] , 通过 作用于预先分泌的 FGF 23 翻译后裂解为活性和非活性片段 [ 96 ]。这种 O-糖基化调节的切割也可能涉及对 ECF Pi 变化的相对缓慢的响应。在临床上,铁状态对慢性肾功能衰竭等常见疾病中的 FGF23 分泌具有重要意义 [ 97 ]。虽然 FGF23 调节 Pi 代谢,PTH 和 1,25D 调节 Pi 和 Ca 代谢,但 Pi ECF 浓度不直接影响 PTH 分泌 [ 98 ],尽管高膳食磷摄入量 [ 98 ],口服 Pi 补充剂 [ 72 ] 间接影响通过降低 ECF Ca 浓度。FGF 23 和 PTH 均通过 NaPi2a 和 NaPi2c 降低 TmPi,但与 FGF 23 降低 1,25D 分泌相比,PTH 增加 1,25D 分泌(图 12 )。因此,这三种激素作为高度整合的激素轴发挥作用,旨在维持 Pi 和 Ca 代谢以及骨矿物质库。此外,这三种激素能够在体外和体内动物模型中调节彼此的分泌。FGF23 减少 PTH 分泌 [ 99 , 100 ],1,25D 减少 PTH 分泌 [ 101 , 102 ] 并增加 FGF 23 分泌 [ 103 ],PTH 增加 FGF 23 分泌 [ 104 ](图 13 )。这些后一种途径在正常生理学和疾病病理生理学中的相对重要性仍然需要一些阐明。应该注意的是,严重的磷酸盐耗竭引起的低磷血症[ [图18 ]与导致高TmPi和1,25D以及低PTH的低FGF23相关。另一方面,由于疾病引起的 FGF 23 分泌增加所致的低磷血症,如 TIO 和 XLH [ 33 ],具有低 TmPi、1,25D 和高正常 PTH。
图 12
ECF磷酸盐浓度[Pi]的增加刺激骨细胞分泌FGF23。增加的 FGF 23 作用于肾小管细胞以减少磷酸盐重吸收 [TmPi] 并减少 ECF Pi,并减少 1,25 二羟基维生素 D [1,25D] 和 Pi 肠道吸收。循环 Pi 浓度 [Pi] 的降低会降低 FGF23 的分泌,从而增加 TmPi 和 125D 的分泌。通过非内分泌机制的低铁状态会增加 FGF 23 的分泌
图 13
FGF 23、PTH 和 1,25D 作为高度整合的激素轴发挥作用,维持 Pi 和 Ca 代谢并调节骨骼中的 Pi 和 Ca 矿物储库
Pi和Ca代谢之间的相互作用
Pi 和 Ca 的代谢不能相互隔离。它们在 ECF、肠道、骨骼和肾脏的运输机制中密切相互作用。此外,Pi 和 Ca 的 ECF 浓度的变化独立地调节维持 Pi 和 Ca 稳态的激素 FGF 23、PTH 和 1,25D 的分泌。在 ECF 中,磷酸八钙过饱和导致软组织矿化,而在饱和下导致骨脱矿。健康的骨形成需要从 ECF 中同时提供必要的 Pi 和 Ca,当骨吸收时,Pi 和 Ca 一起释放到 ECF 中,其量等于它们在磷灰石中的 3/5 摩尔比。在饮食中,Pi 和 Ca 密切相关。乳制品是膳食 Pi 和 Ca 的主要来源。母乳提供约 150 mg Pi 和 180 mg Ca/100 g,与酪蛋白结合为乳胶束,为成长中的婴儿提供高度可吸收的供应。另一方面,饮食中 Pi 的生物利用度会因钙补充剂的大量摄入而显着降低,这种相互作用可用于治疗慢性肾功能衰竭和甲状旁腺功能减退症,以降低 ECF Pi 浓度的增加。相比之下,为治疗低磷血症而摄入的大量 Pi 补充剂会损害钙的吸收,减少循环 ECF Ca,并导致继发性甲状旁腺功能亢进。在肾脏,Ca ECF 浓度增加会降低 TmPi,即使在没有 PTH 的情况下也是如此,并且在尿液中 Ca-Pi 过饱和会增加结石形成的风险。 105 ] 在健康或疾病方面。
不感兴趣
看过了
取消

人点赞

人收藏

打赏
不感兴趣
看过了
取消
©2012-2023 北京华媒康讯信息技术股份有限公司 All Rights Reserved. 注册地址:北京 联系电话:010-82736610
广播电视节目制作经营许可证 —(京)字第 17437号 京海食药监械经营备20200522号
京ICP备12011723号 京ICP证150092号
 京公网安备 11010802020745号
工商备案公示信息
互联网药品信息服务资格证书((京)-非经营性-2020-0015)
京公网安备 11010802020745号
工商备案公示信息
互联网药品信息服务资格证书((京)-非经营性-2020-0015)






您已认证成功,可享专属会员优惠,买1年送3个月!
开通会员,资料、课程、直播、报告等海量内容免费看!

打赏金额
认可我就打赏我~
1元 5元 10元 20元 50元 其它
打赏作者
认可我就打赏我~
扫描二维码
立即打赏给Ta吧!
温馨提示:仅支持微信支付!
已收到您的咨询诉求 我们会尽快联系您












































 010-82736610
010-82736610
 股票代码: 872612
股票代码: 872612




