
SIg,主要是SIgA,通过抑制或促进特定分类群的生长来塑造肠道微生物群。通过直接结合,它们可以影响肠腔中产生的细菌衍生代谢物的数量、组成和位置。
导读
维持共生多样性对宿主体内平衡至关重要,因为微生物物种提供一系列代谢产物,并持续训练宿主免疫系统。粘膜免疫系统必须积极收集有关微生物群组成的信息,同时提供适当的反应。在哺乳动物中,细菌感应导致产生特异性免疫球蛋白(Ig),这些免疫球蛋白以分泌性Ig(SIg)的形式到达肠腔。最近的研究进一步阐明了SIg塑造细菌库并有助于调节宿主代谢的机制。同时,细菌代谢物调节Ig的产生和分泌。本文概述了当前关于细菌代谢物与宿主SIg之间关系的知识,并将这种平衡的破坏与人类慢性炎症联系起来。
论文ID
原名:Metabolites and secretory immunoglobulins: messengers and effectors of the host-microbiota intestinal equilibrium
译名:代谢物和分泌性免疫球蛋白: 宿主-微生物群肠道平衡的信使和效应物
期刊:Trends in Immunology
IF:19.709
发表时间:2021.11
通讯作者:Stéphane Paul
通讯作者单位:法国里昂第一大学
DOI号:10.1016/j.it.2021.11.005
综述目录
1 宿主与其肠道菌群之间的开放对话
2 关于小鼠和人类肠腔中SIg功能的新见解
2.1 SIg可以调节微生物群
2.2 SIg可以抑制微生物群
2.3 SIg支持微生物群
2.4 IgA对细菌代谢物的影响
3 细菌衍生的代谢物可以影响啮齿动物和人类的体液反应
3.1 SCFA可以调节IgA反应
3.2 次级胆汁酸与低IgA分泌有关
3.3 色氨酸衍生代谢物可促进B细胞分化为调节性B细胞
4 破坏细菌代谢物和体液反应之间的平衡:与人类疾病中的慢性炎症有关
4.1 IBD中微生物代谢物的失调
4.2 IBD患者的SIg反应可能增强
4.3 与代谢物/SIgA失调相关的肠外疾病
5 结论
主要内容
1 宿主与其肠道菌群之间的开放对话
肠道是哺乳动物与其环境之间最大的交换表面之一,其主要功能是吸收营养,同时防止病原体进入。为了协调这两种功能,宿主代谢借助于一个高度复杂的生态系统,该生态系统受益于膳食营养素和分泌成分,以交换提供:(i)宿主无法合成的代谢物;(ii)针对病原体定殖的空间和营养竞争;和(iii)直接杀菌活性。然而,这种互惠互动必须受到宿主免疫系统的严格调控,以避免细菌过度生长和维持生态多样性。位于肠道固有层的血浆B细胞(PC)大量产生IgA和IgM,占人类抗体产生量的80%以上。在聚合物Ig受体(pIgR)介导的IgA或IgM通过肠细胞的转胞作用后,这些Ig与pIgR衍生的分泌成分(SC)一起释放到肠腔中。新形成的SIg通过排除过度生长的细菌和积极选择共生体来帮助塑造肠道微生物群。 在小鼠中,至少90%的IgA产生取决于微生物群的存在。微生物多样性通过多种机制感知,包括细菌通过微皱褶(M)细胞转移到派氏集合淋巴结(PPs)以及通过微生物相关分子模式(MAMP)刺激免疫细胞。与MAMP类似,次级胆汁酸(SBA)、色氨酸(Trp)衍生的微生物代谢物和短链脂肪酸(SCFAs)等微生物代谢物可与免疫细胞表达的广谱受体结合,从而调节局部免疫。 在本文中,我们通过解决以下问题来回顾当前对哺乳动物SIg的了解:(i)SIg通过哪些机制塑造微生物群,从而改变细菌衍生代谢物的外显率?(ii)细菌代谢物如何有助于调节免疫球蛋白对微生物群的反应?(iii)代谢产物/SIg平衡的破坏如何参与炎症性肠病(IBD)或其他肠道表现?回答这些问题可能有助于更好地理解肠道慢性炎症的起源和放大机制。
2 关于小鼠和人类肠腔中SIg功能的新见解
SIg可以通过抑制某些分类群和维持其他分类群来塑造肠道微生物群。它们还可以通过M细胞进行反向转胞作用来调节细菌衍生代谢物和腔内样本抗原的分泌(图1和BOX 1)。
图1 分泌性免疫球蛋白(SIg)在小鼠肠腔中的功能。
左图:肠道微生物群的抑制可通过免疫排斥机制介导,如凝集或嵌链生长,以及通过阻断细菌表面蛋白质随后下调其表达。中间:SIg可以通过调节多糖利用位点(PUL)和定殖因子基因来支持肠道微生物群,从而改善细菌适应性和肠道内稳态。SIg还可以通过细菌、免疫球蛋白和粘液相互交织的保护性生态位的形成促进肠道定殖。右图:SIg可能通过增强细菌清除、PUL修饰或目前未知的机制对细菌衍生代谢物产生定性和定量影响。
BOX 1 SIg可促进哺乳动物肠道环境的取样。
对肠腔内容物进行取样可以建立共生体特异性体液反应。微皱褶(M)细胞分布在覆盖PPs和结肠斑块的肠上皮中,专门负责这一功能,与未携带Cre转基因的小鼠相比,缺乏M细胞的Villin-Cre Tnfrsf11af/f小鼠产生的IgA显著减少,粪便中的IgA包被细菌也更少。GP2是沙门氏菌和大肠杆菌表达的1型菌毛受体,其顶端表达允许一些入侵细菌易位。M细胞也能结合SIgA,并允许其从肠腔反向转运至下层淋巴组织。在M细胞的体外模型中,有人提出反向转胞作用是由Dectin-1和Siglec-5的顶端表达介导的;结果证实,缺乏Dectin-1的Clec7a–/–小鼠表现出SIgA介导的反向转胞作用受损。如果SIgA与抗原结合,M细胞与SIgA之间的亲和力会增加,这加强了SIgA介导的反向转胞作用可以帮助取样管腔内容物的想法。FIMH缺陷(invA)沙门氏菌缺乏1型菌毛,因此不与M细胞GP2结合,仅在沙门氏菌先前涂有特异性IgA时在小鼠PPs中观察到,表明IgA允许非侵入性细菌易位至PPs。最近发现了类似的IgM机制;人和小鼠的肠和鼻粘膜M细胞表达TOSO(FcμR),使体外M细胞模型中的SIgM结合抗原易位。经口和鼻腔给药SIgM-HIV-1p24复合物诱导小鼠的免疫反应比单独给药p24更强。 SIgA还被怀疑能够保护完整的醇溶蛋白肽从管腔运输到固有层,这是腹腔疾病病理生理学中的一个关键步骤。活动性乳糜泻患者的十二指肠上皮顶端异常表达CD71,在该病患者的十二指肠活检中观察到IgA结合醇溶蛋白的反向转胞作用,当联合施用抗CD71抗体时,该现象被消除。
2.1 SIg可以调节微生物群
许多小鼠模型证实了分泌性IgA(SIgA)在维持健康肠道微生物群中的重要性。缺乏激活诱导胞苷脱氨酶(Aicda–/–)(B细胞进行类别转换重组(CSR)和体细胞超突变(SHM)所必需的酶)的小鼠,在小肠中大量积聚大量厌氧微生物。与野生型(WT)小鼠相比,缺乏SIgA和SIgM的Pigr–/–小鼠表现出肠道上皮屏障的显著降解,这反映在血清蛋白泄漏和粪便白蛋白浓度升高,盲肠微生物多样性较低,对葡聚糖硫酸钠(DSS)诱导的结肠炎的易感性较高。与野生型同窝小鼠相比,具有选择性IgA缺乏症(IghA–/–)的小鼠的肠道微生物群多样性也较低,这表明IgA在调节肠道微生物群和内稳态方面起着重要作用。 在人类中,选择性IgA缺乏症(SIgAD)患者队列中也有轻度肠道生态失调的报道,SIgAD是一种常见的免疫疾病,其发病机制未知。IgM可能部分补偿了SIgAD,因为与健康个体相比,SIgAD患者粪便中的IgM浓度和IgM包被细菌比例更高。事实上,来自SIgAD患者的IgM包被细菌与健康个体的IgA包被细菌存在显著差异,表明IgA和IgM的微生物结合模式仅部分重叠,并且IgM的代偿作用在一定程度上受到限制。患有常见可变免疫缺陷的患者也表现出肠道生态失调,与健康对照组相比,与放线菌减少相关的Gammaproteobacteria显著增加,这表明免疫球蛋白对于维持健康的肠道微生物群至关重要。
2.2 SIg可以抑制微生物群
在肠腔中,SIgA可包裹并凝集细菌,导致形成团块,然后通过消化和/或蠕动消除团块,该过程称为免疫排斥。最近,一种被称为“enchainedgrowth”的细菌凝集机制被发现。具体而言,在感染鼠伤寒沙门氏菌的小鼠中,IgA结合阻止快速分裂的细菌在分裂后分离,导致子细胞被迫以“enchained”方式生长,并形成更易于被肠蠕动流消除的单克隆团块。 IgA还可以通过结合特定的表面蛋白来抑制微生物群。例如,脂多糖(LPS)特异性Sal4单克隆IgA与鼠伤寒沙门氏菌的结合可显著降低体外细菌的运动性。此外,据报道,从无菌WT小鼠的肠道PC产生的W27 IgA单克隆抗体(mAb)可特异性抑制大肠杆菌的丝氨酸羟甲基转移酶(SHMT),从而限制其体外生长。此外,用W27单克隆抗体对小鼠进行口服治疗可减少微生物失调和随后DSS诱导的结肠炎严重程度,从而支持SHMT结合在调节体内微生物多样性中的潜在作用。值得注意的是,当与鞭毛蛋白特异性IgA共培养时,共生菌如大肠杆菌、Roseburia intestinalis或Clostridium ramosum的运动也可以在体外受到抑制;据报道,这不仅是由鞭毛蛋白的直接空间位阻引起的,而且是由结合诱导的鞭毛蛋白基因表达下调引起的,如使用GFP融合鞭毛蛋白基因构建的大肠杆菌菌株所示。IgA结合介导的基因调节也在单殖Bacteroides thetaiotaomicron的Rag1–/–小鼠中观察到,并接种了针对荚膜多糖的单克隆IgA(225.4)。与225.4结合可诱导靶向多糖表位的下调,降低一氧化氮代谢相关基因的表达,降低B. theta肠道密度;这表明IgA通过影响基因表达来影响细菌适应性。值得注意的是,已提出IgA结合可调节细菌代谢活动,如下所述。
2.3 SIg支持微生物群
SIgAD患者的轻度肠道失调的特征不仅是致病菌增加,而且共生菌减少,这表明IgA支持共生细菌生长。事实上,在小鼠身上的实验表明,IgA有助于细菌定殖。具体而言,单定殖Bacteroides fragilis的WT小鼠表现出粘液和肠粘膜中缠绕的细菌聚集体,这在Igha–/–小鼠中是不存在的。此外,在B. fragilis和HT29上皮细胞的共培养物中添加B. fragilis特异性IgA,但不是无关的IgA,导致细菌与上皮细胞的粘附增强,支持IgA在B. fragilis肠道定殖中的作用。与对照组相比,将一种糖基化卵清蛋白特异性单克隆IgA(7-6IgA)施用于有B. theta定殖的Rag1–/–小鼠,不仅改善了B. theta的适应性,而且还导致梭菌的增殖和对DSS结肠炎的抗性增加。除了细菌组外,IgA还可以与肠道真菌群落相互作用,以支持肠道内稳态。具体而言,在单定殖白色念珠菌(Candida albicans)的WT小鼠中,成像流式细胞术突出显示白色念珠菌IgA+部分菌丝显著富集;用肠道IgA探测酵母和菌丝细胞壁组分,发现其优先结合菌丝组分。此外,液相色谱和质谱分析表明,粘附素是IgA靶向菌丝部分最丰富的蛋白质,表明IgA与粘附素结合抑制白色念珠菌菌丝形态,同时促进酵母形式,从而维持肠道内稳态。 目前,对SIg与人类病毒组之间的关系知之甚少。SIgAD和CVID患者口腔病毒组的宏基因组分析显示,与健康对照组相比,唾液病毒组组成和/或多样性没有显著差异,表明IgA对维持病毒组内稳态并不重要,但这仍有待进一步评估。因此,最近在单定殖小鼠模型和人类身上的研究正在修正IgA的功能,并强烈表明SIgA在某些肠道共生体的生长中起支持作用。
2.4 IgA对细菌代谢物的影响
为了研究Ig在细菌代谢物渗透宿主生物体中的作用,将13C标记的非复制大肠杆菌注射到缺乏抗体的无菌WT和Igh-J–/–小鼠体内。Igh-J–/–小鼠的血清和肠液中13C标记代谢物浓度增加,而WT小鼠的肠道转运和粪便细菌脱落速度加快。此外,从WT小鼠中分离出的大肠杆菌表明,IgA与鞭毛的结合影响了细菌的运动,因此限制了细菌繁殖和代谢物进入宿主肠道。通过研究MD4小鼠的微生物群(其抗体库仅限于单一非细菌抗原),发现能够将L-酪氨酸转化为硫酸对甲酚(PCS)的细菌富集,并假设其归因于异常IgA反应。
IgA还可以通过调节细菌基因表达影响肠道代谢组。来自与B. theta单定殖的WT小鼠的多克隆IgA主要结合来自多糖利用位点(PUL)的蛋白质(细菌利用果聚糖所必需)。细菌与IgA结合后,这些PUL蛋白的表达下调,表明IgA通过阻碍多糖的利用来限制细菌代谢。相比之下,一种新鉴定的B. theta PUL,命名为MAFF,在体外通过聚糖-聚糖相互作用与非特异性单克隆IgA结合后表达增加。用WT B. theta菌株(而不是缺乏MAFF的菌株)对小鼠进行定殖,可诱导产生丁酸的梭菌增殖,这与对DSS诱导的结肠炎的保护有关。此外,在与B. theta单定殖的小鼠中产生的肠道多克隆IgA也被发现结合并抑制果聚糖PUL,影响果聚糖衍生的代谢组。 最近的两份报告强调了IgA包被的乳酸杆菌与代谢之间的联系。在第一项研究中,与正常饮食的小鼠相比,喂食高脂肪饮食的小鼠结肠中SCFA的生成显著减少。当从健康个体的粪便中接种IgA包被的乳酸杆菌(而不是单独的乳酸菌)时,SCFA产量的下降部分得到缓解。为什么只有IgA包被的乳酸杆菌才能缓解表型仍有待解决。另一项研究使用P2rx7–/–小鼠,其缺乏ATP门控离子受体P2X7,因此表现出代谢紊乱;这些小鼠的特征是PPs中的滤泡辅助性T细胞(Tfh)增加。与野生型同窝小鼠相比,在这些小鼠中观察到肠道生态失调和肠道细菌(尤其是乳杆菌)IgA涂层增强,这表明Tfh扩增调节了IgA对微生物群的反应。此外,用P2rx7–/–小鼠IgA包被乳酸杆菌灌胃的WT小鼠表现出葡萄糖稳态和脂肪沉积的改变,这表明在P2rx7–/–小鼠中观察到的代谢紊乱在某种程度上与IgA包被乳酸杆菌的积累有关。因此,本研究将细菌的IgA涂层与宿主代谢缺陷联系起来;然而,细菌代谢物在这一过程中的作用仍然难以确定。 总之,这些结果表明,共生体的IgA涂层通过调节微生物群衍生的代谢物积极或消极地改变宿主代谢稳态。假设这些作用是独特的,可能取决于细菌种类、粘膜免疫细胞激活状态以及IgA靶点。微生物群衍生的代谢物也可能反过来影响宿主的体液免疫反应。
3 细菌衍生的代谢物可以影响啮齿动物和人类的体液反应 3.1 SCFA可以调节IgA反应
肠道微生物群从其他不可消化的膳食纤维中产生SCFA,不仅为宿主提供代谢供应,而且有助于免疫系统的调节。早期研究表明,与对照组相比,喂食膳食纤维的大鼠盲肠分泌更多的IgA,盲肠和小肠粘膜中有更多的IgA+ B细胞,以及肠系膜淋巴结(MLN)。这在喂食高膳食纤维或醋酸盐的抗生素治疗后的老鼠身上得到了证实。有人提出,SCFA可直接促进PC代谢,因为给小鼠分离的脾B细胞注入乙酸盐、丁酸盐或丙酸盐可增加线粒体呼吸和随后的ATP/ADP比率。在第二项研究中,未观察到乙酸对纯化的脾脏B细胞有直接影响,但与经醋酸盐预处理的纯化的树突状细胞(DCs)共培养的B细胞增加了DC维甲酸(RA)和随后的B细胞IgA的产生,这可以用RA受体拮抗剂消除。因此,醋酸盐被提议作为IgA产生的促进剂,提高DC将维生素A转化为RA的能力。然而,在略有不同的研究中观察到相反的结果。喂食18%纤维饮食的小鼠,MLNs、PPs和脾脏中的IgA+细胞较少,粪便中的IgA包被细菌较少,而IgM的产生量仍然较低。喂食高剂量丁酸盐和丙酸盐的小鼠再现了先前的观察结果,即肠道中产生的IgA较少,并且在用NP-CGG或NP-LPS免疫后,小鼠呈现NP3特异性IgM和IgG滴度降低,与对照小鼠相比SHM比率较低。作者得出结论,丁酸和丙酸抑制SHM。用纯化的人或小鼠B细胞进行的体外实验证实,在激活后,以及在存在高浓度丁酸和/或丙酸盐的情况下,CSR和SHM降低。 与对照组相比,食用富含醋酸盐饮食的无菌非肥胖糖尿病(NOD)小鼠盲肠中的IgA分泌减少,微生物群的IgA涂层减少,PPs中的生发中心显著减少。这一观察结果与醋酸盐处理后肠道pIgR表达的降低相关。此外,与仅用LPS和抗CD40单抗刺激的B细胞相比,从NOD小鼠纯化的B细胞的LPS、抗CD40单抗和醋酸盐刺激也减少了与PC分化(Psta、Pst2b、Stat5b和Irf4)和IL-6产生相关的转录物数量;这表明醋酸盐对NOD小鼠B细胞有直接影响。值得注意的是,与健康对照组相比,1型糖尿病(T1DM)患者的粪便IgA结合细菌增多,粪便SCFA浓度降低,这两个参数在T1DM患者中呈负相关,说明醋酸盐对IgA生成的潜在调节作用。 此外,当与大肠杆菌单定殖时,口服补充了醋酸盐的小鼠在大肠中表现出IgA生成增加,但与B. theta单定殖时没有。因此,体外实验证实,当补充大肠杆菌LPS和醋酸钠时,与来自WT小鼠的CD4+ T细胞共培养的B细胞(而不是来自缺乏TLR信号的Myd88–/–Ticam1–/–小鼠的B细胞)显示出IgA生成增加。醋酸盐可能有助于以TLR信号相关的方式刺激CD4+ T细胞反应,导致PC产生更多IgA,但这仍有待严格测试。总之,这些研究表明,SCFAs对IgA的产生具有不同的、有时是相反的影响,我们认为这可能取决于SCFA的数量、微生物群组成和遗传背景。已经提出了SCFA对B细胞的一些直接作用(见上文),以及可能涉及DC或CD4+ T细胞的间接作用。然而,这些假设需要进一步稳健的测试。
3.2 次级胆汁酸与低IgA分泌有关
初级胆汁酸(PBA)在肝脏中合成,分泌到胆管,并释放到十二指肠腔。大多数PBA在回肠中被重新吸收,并通过门静脉再循环到肝脏。然而,约5%的PBAs逃逸循环并到达结肠,在结肠中,它们被常驻细菌代谢生成SBA。SBAs抑制小鼠巨噬细胞的NLRP3炎性小体激活,与未经治疗的对照组相比,石胆酸衍生物可降低小鼠辅助性T细胞17(Th17)/调节性T细胞(Treg)比率;这表明一些SBA具有抗炎特性。此外,对人外周血单个核细胞(PBMC)的早期研究表明,在金黄色葡萄球菌Cowan I体外刺激下,SBA熊去氧胆酸对Ig产生负调控。此外,与对照组相比,喂食添加PBA胆酸的饮食的大鼠回肠粘膜中IgA的生成显著增加;这表明,与SBA相比,PBA增加了IgA的产量。然而,最近没有研究证实SBAs对IgA分泌的影响,还需要进一步研究。然而,从小鼠脾脏纯化的B细胞表达少量编码SBA受体的Tgr5转录本,表明SBA影响B细胞活性。总的来说,需要进行研究以确定某些SBA分子是否参与假定的IgA生成调控,以及这是否可能是物种或环境特异性的。
3.3 色氨酸衍生代谢物可促进B细胞分化为调节性B细胞
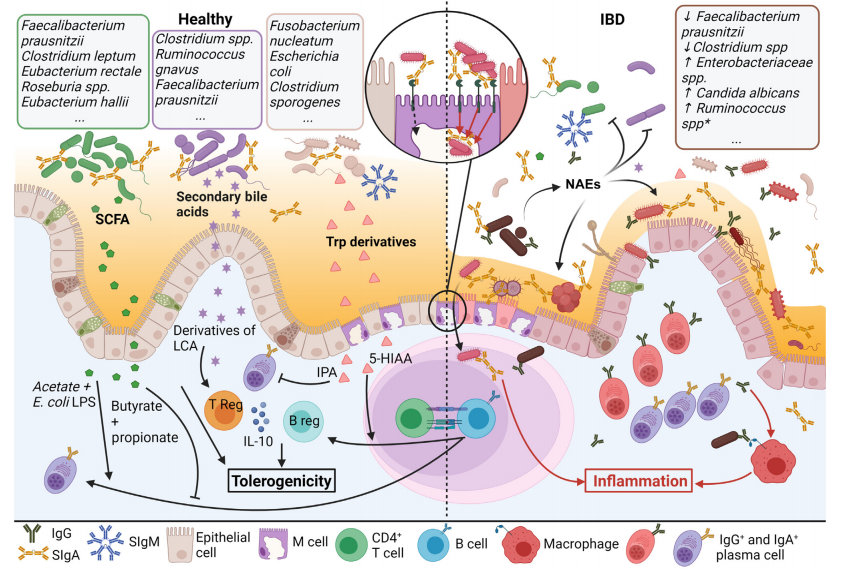
芳香烃受体(AhR)参与广泛免疫细胞的调节。AhR在B细胞中的表达在激活后增加,其在小鼠中的参与抑制了细胞向PCs的分化,并在体外和体内分泌IgA和IgM,有利于B细胞分化为调节性B细胞(Breg)。多种色氨酸(Trp)衍生代谢物是AhR配体,包括吲哚衍生物,由肠道微生物群的特定分类群代谢。事实上,据报道,与WT菌株单定植的小鼠相比,不能将色氨酸代谢为吲哚丙酸(IPA)的Clostridium sporogenes(Cs)菌株单定植的无菌小鼠在盲肠中分泌更多Cs特异性IgA,表明细菌Trp代谢减少了特异性IgA的分泌。因此,与对照组相比,喂食缺乏AhR配体饮食的小鼠产生的粪便IgA较少,除非补充吲哚-3-甲醇。此外,在抗原诱导的关节炎小鼠模型中,口服丁酸盐补充物富集了Trp代谢细菌,导致粪便中5-羟基吲哚-3-乙酸(5-HIAA)的增加(相对于正常饮食喂养的小鼠)。与对照组相比,用5-HIAA体外处理脾脏B细胞可增加Il10的表达,并且与对照组灌胃小鼠相比,5-HIAA灌胃小鼠脾脏B细胞的Il10表达也增加。除非保持无菌状态,否则Il10–/–小鼠会自发地发展成结肠炎,并且其粪便中的IgA和IgG滴度高于野生型同窝小鼠。因此,IL-10被怀疑对抑制肠内IgA产生和向IgG产生的类别转换很重要,尽管这仍有待证明。综上所述,虽然微生物代谢促进或抑制IgA产生的机制尚未完全阐明,但越来越清楚的是,IgA的产生和微生物代谢是相互关联的(图2)。这种平衡的丧失与慢性肠道炎症,甚至肠外表现密切相关。因此,作为这些代谢物主要来源的饮食质量也应考虑在内,以便更好地了解IgA产生和宿主微生物群对肠道内稳态的影响。
图2. 细菌衍生代谢物和免疫球蛋白(Ig)反应可在体内平衡期间调节炎症,在炎症性肠病(IBD)中调节失调。
在健康的微生物群中,分泌性Ig(Sig)通过共生体促进粘液定殖,排除了快速分裂的细菌,并通过Sig与微皱褶(M)细胞结合采集抗原。这种永久性的选择压力有助于维持具有一系列代谢活动的高度多样化的微生物群。新产生的代谢物可能有助于其他类群的代谢,或被上皮细胞吸收到固有层。在这些代谢物中,短链脂肪酸(SCFA)、次级胆汁酸(SBA)和色氨酸(Trp)衍生物是体液肠道免疫的重要调节因子,可通过小鼠和/或人体内发现的不同途径调节IgA的产生。产生这些代谢物的已知分类群示例见上框。IBD患者的粘膜特征是Ig产生显著增加,尤其是IgA和IgG。在它们的肠腔中,一些次级代谢物被富集,支持致病菌的生长,同时抑制共生菌的生长。右上框显示了IBD患者粪便中分类群丰度的主要变化,星号表示伴随IgA涂层的增加。通过M细胞增加SIgA逆转录转运也可能通过促进细菌易位而促进炎症反应。固有层中的抗原结合IgG可能从肠腔再循环,也可通过FcγR结合激活巨噬细胞炎性小体途径参与局部炎症。平头箭头表示负调节,尖头箭头表示正调节。缩写:5-HIAA,5-羟基吲哚乙酸;Breg,调节性B细胞;IPA,3-吲哚丙酸;LCA,石胆酸;LPS,脂多糖;NAEs,N-酰基乙醇胺;Treg,调节性T细胞。
4 破坏细菌代谢物和体液反应之间的平衡:与人类疾病中的慢性炎症有关
4.1 IBD中微生物代谢物的失调
IBD分为两组,克罗恩病(CD)和溃疡性结肠炎(UC),均以慢性胃肠道炎症为特征。20多年来,这两种形式都与人类的肠道失调有关。同样,如果在无菌环境中饲养,用于自发性结肠炎模型的大鼠也会受到保护。IBD中的微生物代谢物高度失调,这与生态失调一致。鉴于这些改变已在其他地方得到广泛研究,我们在此重点讨论经实验证实的代谢物失调(表1),以及确认或重申先前观察结果的体内小鼠实验的细节。与前面章节的讨论相关,IBD患者粪便中SCFA(丁酸盐和丙酸盐)、吲哚衍生物和SBA的含量减少。在特定的小鼠模型中,这些代谢物的缺失会加重实验性结肠炎,而补充这些代谢物会降低结肠炎的严重程度(表1)。代谢物结合途径也很有趣:从CD患者或对照患者粪便中纯化的微生物群被转移到Il10–/–无菌小鼠,旨在放大肠道共生致病菌。因此,在感染CD患者微生物群的无菌Il10–/–小鼠中,几种硫酸化化合物显著富集,包括硫酸化胆汁酸。鉴于硫化氢(H2S)被视为在IBD中起着有害的作用,作者认为硫酸盐细菌代谢物的去结合有助于疾病的进展,值得进一步关注。
表1. IBD患者粪便中细菌衍生代谢物失调的例子。
a.↑和↓表明与健康患者相比,仅CD患者或CD和UC(IBD)患者粪便中的代谢物丰度分别增加或减少。缩写:Tgr5,G蛋白偶联胆汁酸受体(也称为GPBAR1)。
4.2 IBD患者的SIg反应可能增强
IBD患者粪便中的总Ig含量较高,炎症粘膜中的PCs含量增加。事实上,与健康对照组相比,IBD患者粪便中的IgG-、IgM-和IgA结合细菌显著增加,高IgG和IgA细菌包被与CD活性相关。此外,SIgAD是人类IBD的一个风险因素,Igha–/–小鼠表现为加重的结肠炎,加强了IgA在形成多样化微生物群中的重要性,从而限制了IBD的发生。然而,一些观察结果也表明增加免疫球蛋白的产生在维持炎症状态方面起着有害作用。首先,与抗体介导免疫选择(AMIS)的概念一致,IgA可促进肠道内特定细菌菌株的定殖。在IBD中,如IgA包被细菌的16S测序所示,SIgA特异性结合到致病生物。当转移到无菌小鼠时,与来自同一供体的IgA-细菌相比,IgA+细菌定殖于受体小鼠正常无菌的内粘液层并恶化DSS结肠炎。IBD患者的IgA是否优先与能够定殖内粘液层的细菌结合,或IgA结合是否促进这些致病菌的定殖,尚待确定。其次,30–50%患有CD的高加索人携带NOD2突变,与M细胞中Dectin-1和Siglec-5表达增加相关,从而相对于健康患者或未携带NOD2突变的患者加速IgA向PPs的逆转录转运。因此,与野生型同窝小鼠相比,Nod2–/–小鼠表现出IgA结合的HIV-1亚单位p24易位增加,但不单是p24易位增加;与对照组相比,前者还显示粪便TNF-α、IFN-γ和IL-17浓度增加,表明IgA介导的抗原逆转录转运参与了局部炎症。IBD患者血清中SIgA滴度升高也可能反映了SIgA转运增加和/或上皮通透性增加,因为血清SIgA仅来源于粘膜分泌物。 第三,如上所述,IBD患者的粪便结合和细菌结合IgG滴度异常升高,反映了其肠腔中的IgG分泌。IgG-抗原复合物是FcγR信号的有效诱导物,管腔IgG-抗原复合物也可能通过与新生儿Fc受体(FcRn)结合而循环回肠固有层。抗鞭毛蛋白IgG(而非对照IgG)在小鼠静脉注射时加重DSS结肠炎。Fcrn–/–小鼠给予抗鞭毛蛋白IgG后可防止结肠炎恶化,这表明鞭毛蛋白-IgG复合物的逆转录转运有助于粘膜炎症。与这一假设一致,导致FcγR信号增加的FCGR2A变体与UC有关,与同窝野生型小鼠相比,显示FcγR信号增加的Fcgr2b–/–小鼠对DSS结肠炎的反应更为严重。总之,这些观察结果表明,IgG和SIgA逆转录转运在IBD中可能起到有害作用,尽管这仍有待深入研究。
4.3 与代谢物/SIgA失调相关的肠外疾病
患有CD和周围脊柱关节炎并发症的患者的粪便中表现出IgA结合的大肠杆菌菌株富集。与未定殖的同窝小鼠相比,这种细菌对Il10–/–或K/BxN小鼠的定殖分别加重了它们的结肠炎或炎性关节炎;这支持了生态失调导致肠道和关节炎症的概念,尽管这需要进一步研究。与无肠外表现的CD患者相比,所有形式的脊椎关节炎患者的血清中也发现了来自粘膜分泌物的SIgA滴度更高。其他肠外表现也与微生物群和免疫系统之间失去平衡有关。最后,肠道异常IgA反应与微生物代谢组受损有关,并在自身免疫性川崎病(KD)、IgA肾病、肌萎缩侧索硬化症(AML)、多发性硬化症(MS)、阿尔茨海默病(AD)、自闭症谱系障碍(ASD)、精神分裂症、慢性抑郁症、慢性疲劳综合征(表2)患者中得到确认。这些报告大多集中在SIgA上,因为它是肠内产生的最丰富的同型。然而,更准确地记录肠外表现中肠道IgM和IgG的调节也很重要。事实上,肠道微环境及其与体液免疫的联系需要在广泛的疾病中加以考虑,这可能是未来研究的一个富有成果的领域。
表2. 人类肠外疾病的粘膜起源,通过改变的IgA滴度和肠道衍生代谢物的组成揭示。
结论
SIg,主要是SIgA,通过抑制或促进特定分类群的生长来塑造肠道微生物群。通过直接结合,它们可以影响肠腔中产生的细菌衍生代谢物的数量、组成和位置。反过来,这些代谢物可以调节粘膜体液反应。SIg和细菌代谢物之间的平衡受到严格调控,其破坏与肠道和肠外慢性炎症有关。然而,其潜在机制仍有待研究,以揭示肠粘膜的生理机能,更好地理解肠道和肠外慢性炎症性疾病的病理生理学。更好地理解宿主免疫系统和肠道微生物群之间的关系可能有助于将细菌代谢物识别为具有诊断和预后意义的假定生物标志物和/或潜在地作为治疗某些慢性炎症性肠道疾病的候选治疗靶点。此外,环境因素(如营养)对细菌代谢组和粘膜免疫反应的影响也有待进一步研究。每个个体都有一个独特的微生物群/Ig特征,破译这种相互关联的生物学特性有助于进一步为个性化医疗铺平道路。
原文链接:https://www.cell.com/trends/immunology/fulltext/S1471-4906(21)00241-6
编译:微科盟听雪斋,编辑:微科盟居居、江舜尧。
微科盟原创微文,欢迎转发转载,转载须注明来源《微生态》公众号。
不感兴趣
看过了
取消

人点赞

人收藏

打赏
不感兴趣
看过了
取消
©2012-2023 北京华媒康讯信息技术股份有限公司 All Rights Reserved. 注册地址:北京 联系电话:010-82736610
广播电视节目制作经营许可证 —(京)字第 17437号 京海食药监械经营备20200522号
京ICP备12011723号 京ICP证150092号
 京公网安备 11010802020745号
工商备案公示信息
互联网药品信息服务资格证书((京)-非经营性-2020-0015)
京公网安备 11010802020745号
工商备案公示信息
互联网药品信息服务资格证书((京)-非经营性-2020-0015)






您已认证成功,可享专属会员优惠,买1年送3个月!
开通会员,资料、课程、直播、报告等海量内容免费看!

打赏金额
认可我就打赏我~
1元 5元 10元 20元 50元 其它
打赏作者
认可我就打赏我~
扫描二维码
立即打赏给Ta吧!
温馨提示:仅支持微信支付!
已收到您的咨询诉求 我们会尽快联系您



































 010-82736610
010-82736610
 股票代码: 872612
股票代码: 872612




