
转移是造成癌症致死的主要原因,LncRNA 不仅是癌症发生发展中重要的调节因子,而且是转移发病机制中的关键因素。
转移是癌症主要的致死原因。它是一个多步骤过程,原发肿瘤细胞通过分子和表型变化得以在远处器官转移定植。过去的研究发现了长链非编码 RNA (lncRNA), 并揭示了它们调节作用的多样性,包括其在肿瘤转移过程中的关键作用。lncRNA 通过影响转移前步骤(包括上皮-间质转化、侵袭和迁移以及器官定植)以及转移性肿瘤微环境来促进转移。本文讨论了体内特征性 lncRNA,以及它们在机制上的共性,例如其与TGFβ-ZEB1/ZEB2 轴或核因子-κB通路的融合,同时对lncRNA目前仍具有争议的机制进行解释。此外,一些 lncRNA 可帮助识别高转移风险的肿瘤并指导治疗。其中一些 lncRNA 在动物模型中可作为基因治疗的潜在靶点。除了特征性 lncRNA 在肿瘤转移中的作用外,本文还讨论了目前lncRNA研究的争议和新的挑战,并为这个快速发展的领域提出了未来研究的方向。
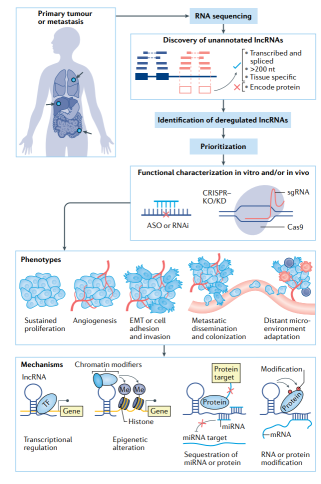
转移是癌症主要的致死原因。虽然癌症的诊断和治疗取得了长足的进展,转移性癌症患者的预后仍然很差,某些癌症中位生存时间仍以月为单位。肿瘤细胞从原发肿瘤扩散至远处器官被称为侵袭-转移级联反应 。原发性肿瘤细胞必须局部侵入周围的正常组织,然后,这些细胞渗入体循环并随后在远处定植。 长非编码RNA(lncRNAs)定义为超过200个核苷酸的RNA转录本,没有证据表明它们能够编码肽链(图1)。lncRNAs最初仅被描述为基因转录子,包括反义转录子、重复转录子和内含子等,其功能取决于LncRNA基因组邻近的蛋白质编码基因。高通量技术已破解数万个LncRNAs序列,但只有一小部分的功能被发现。通过对癌症标本的表达分析和转录组学比较,将优先研究各种LncRNAs功能,并揭示其表型和机制(图1)。
LncRNAs在基因调控中发挥了重要作用,包括调节基因的激活、沉默,X染色体失活,选择性剪切和翻译后调控。LncRNAs通过多种不同的机制发挥这些功能,如在HOTAIR或DLX6AS中,充当“引导”染色质的分子支架-修饰酶,作为“海绵状”microRNA或蛋白质的竞争性内源性RNA(ceRNAs),促进或抑制长链染色质相互作用(例如,LUNAR1或CCAT1),甚至通过转录本身发挥作用(图1)。其他机制也参与作用,比如协调核结构,形成环状LncRNAs使相互作用的mRNAs的不稳定。特定ceRNA模型的机制引起了广泛关注,主要是由于靶mRNA与其在ceRNAs上结合位点之间的化学计量不平衡。由于一些LncRNAs具有多种作用机制,导致该研究领域的复杂性。
LncRNA 是肿瘤发病通路的关键调节因子,也是肿瘤发展的生物标志物 。LncRNA最初是分子生物学领域的一个概念,现在已被发现作用于大多数肿瘤病程的经典通路——持续增殖、复制永生、逃避生长抑制因子、诱导血管生成、抗细胞凋亡和转移(图1).重要的是,就像蛋白质编码基因,遗传改变是肿瘤发生的基础,LncRNA 在恶性肿瘤中扩增、清除或突变。LncRNA 广泛位于肿瘤基因组,包括 FAL1基因,其扩增能够抑制生长抑制基因 CDKN1A。PVT1 在乳腺癌和其他肿瘤中也异常扩增,在其启动子区域中发现了反复突变。在肿瘤中,LncRNA 也发生了的表观遗传学改变,如CpG 甲基化表型中的 CCAT1 激活。因此,LncRNA 的遗传和表观遗传改变可能是早期癌症基因组研究导致罕见突变的原因。
与蛋白质编码基因相反,蛋白质编码基因的结构域决定了其功能。LncRNA 作用机制种类繁多,而且仍在不断涌现。PVT1 具有作为 DNA 调节元件(即顺式作用元件)和 RNA 转录物(即反式作用因子)的功能。这并不是 PVT1 特有的,需要建立适当的模型来分析DNA 或 RNA对肿瘤发生和转移的作用。随着 LncRNA 肿瘤生物学领域研究的进展,研究 LncRNA 的独立研究小组使用不同的研究方法导致不少结果存在矛盾。虽然这些相互矛盾的结论存在争议,也揭示了 LncRNA 的复杂性,并促使了研究方法的改进。
我们对LncRNAs如何调节侵袭转移级联反应以及其器官特异性,对肿瘤微环境(TME)的影响的认识正在稳步增加(图2,3)。已有研究致力于LncRNA在癌症转移中的作用,但最近的几项研究质疑了之前关于LncRNA功能和机制的研究。重要的是,如改良反义核苷酸(ASOs)和CRISPR–Cas9及其衍生物改变了LncRNAs的研究方式,对于LncRNA功能的研究有了新的启示。在这篇综述中,我们将讨论LncRNA在癌症转移中的作用,以及与LncRNA有关的争议。
图1 肿瘤中的LncRNA 局部侵袭和EMT
周围组织局部侵袭是肿瘤转移的必要初始步骤。上皮-间质转化 (EMT) 及其反向间质-上皮转化是在胚胎发育和组织稳态过程中被正常激活以确保组织和器官正常形态。对于侵袭-转移级联反应至关重要。通过激活主要调节因子如 SNAI1、TWIST1 和 ZEB1,具有上皮表型的癌症细胞被赋予间充质细胞的特征,具有侵袭、血管内渗入和远处转移的功能。尽管许多 LncRNA被认为可以调控生长和侵袭,我们将讨论重点放在在侵袭和 EMT 中发挥作用的 LncRNA,这些LncRNA 的作用在体内转移实验中得到证实(图 2)。

图2 LncRNAs与侵袭-转移级联反应
EMT 中的 H19
H19 是最早被研究的LncRNA 之一,在多种癌症中过度表达并参与肿瘤细胞侵袭(图 3)。在膀胱癌细胞中,H19 过表达导致了肿瘤细胞在体外转移,与多梳抑制复合物2(PRC2)成分EZH2相作用,导致β-连环蛋白活性升高和E-钙粘蛋白活性降低,向间充质状态过渡。目前争议焦点在于PRC2有与RNA相互作用的趋势,这可能会与发挥作用的H19主要分子相混肴。H19 介导的间充质转化也发生在结直肠癌 (CRC) 细胞中,H19是一种ceRNA,阻断microRNA,miR-138和miR-200a,导致波形蛋白、ZEB1和ZEB2蛋白表达的降低。H19在间充质亚型结直肠癌细胞中的表达始终高于上皮亚型。H19促侵袭并最终导致转移的作用在自发性乳腺肿瘤小鼠模型中得到进一步支持。H19在能够定植转移的细胞克隆株中过表达(即4T1、4T07和168FARN株系),由短发夹RNA(shRNA)的介导的H19基因敲除可消除从原发部位到远端器官的转移(如肺,肾脏和肝脏)。在这项研究中,H19作为针对miR-200b 和 miR-200c的ceRNA,导致原发性肿瘤和远端转移瘤中 ZEB1 和 GIT2 的抑制。GIT2可以促进远处转移和定植。在循环肿瘤细胞(CTCs)中,H19差异地粘附miRNA let-7b,导致CYTH3抑制,有利于促进间充质表型外渗。这突出了环境背景对LncRNA功能的显著影响,是LncRNA一般生物学特性。因此,基于潜在细胞和环境背景不同作用机制,H19在转移起始和定植级联反应中起到了协调的作用。
目前对于H19功能的研究并未区别H19转录物microRNA的作用与其自身产生miR-675的作用。后者编码在H19基因的第一个外显子中。在前列腺癌细胞中,H19高表达与转移潜能降低相关,并且H19编码的mir-675表达导致转化生长因子 β 诱导蛋白 (TGFBI) 下调,TGFBI 参与 EMT的进展。因此,源自H19基因转录物可能具有分离调节侵袭转移级联的作用。尽管将单一调节功能归因于LncRNA更简单,有理由认为,在多种癌症类型中LncRNA (例如H19) 可能具有更关键的作用和多种调节机制。

图3 LncRNAs调节转移的机制
EMT 中的其他竞争性内源性 LncRNA
LncRNA与EMT有关。其中,促过渡相关 RNA (PTAR) 与卵巢癌的间充质亚型相关,并作为针对 miR-101的ceRNA,导致 ZEB1 表达的抑制。shRNA 介导的 PTAR 敲除卵巢癌异种移植物中 E-钙粘蛋白表达增加并降低了 FN1、ZEB1 和波形蛋白的表达。此外,与对照组相比,腹膜内注射PTAR 敲除卵巢癌细胞的小鼠的肿瘤结节更少,表明了PTAR 在促进 EMT中的作用。 LncRNA-PNUTS,是从产生 LncRNA 变体 (LncRNA-PNUTS) 和 mRNA 变体 (PNUTS) 的基因座转录而来。虽然 LncRNA作为ceRNA,但mRNA产生蛋白磷酸酶1调节剂。使用 TGFβ 处理细胞可导致PI3K-Akt 依赖性磷酸化LncRNA-PNUTS 转录抑制。LncRNA-PNUTS 表达与 ZEB1 和/或 ZEB2 间充质标志物表达相关,表现为与 miR-205 结合的 ceRNA形式,导致 ZEB1/ZEB2 的表达抑制(图 4)。在肺癌和小鼠乳腺细胞系中,LncRNA-PNUTS的过表达可诱导 EMT,这可以通过 ZEB1/ZEB2 和波形蛋白的形态改变和表达增加来实现,这种效应可以通过与 miR-205 共转染而被消除。ShRNA 介导的LncRNA-PNUTS 敲除MDA-231-LM2 乳腺癌细胞系注射到小鼠乳腺脂肪垫后,肺转移的负担减少。
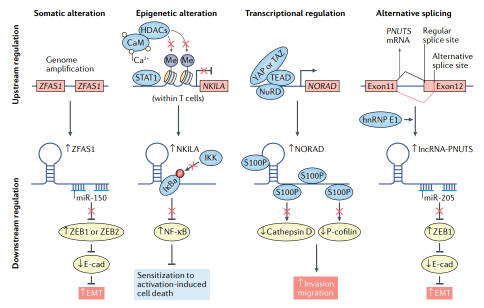
由 TGFβ 激活的 LncRNA (LncRNA-ATB) 是另一种 ceRNA,作为 TGFβ-ZEB1/ZEB2 轴的调节剂。作为 TGFβ 的转录靶标,LncRNA-ATB 竞争性结合 miR-200,导致 ZEB1/ZEB2 表达上调。与该机制一致,miR-200a 在肝癌细胞中的过表达消除了 EMT。因此,LncRNA-ATB 在肝细胞癌 (HCC) 的体内原位异种移植模型中可以诱导 EMT、侵袭、增加CTC 水平和远处器官定植。同样,在 HCC 细胞中,lncRNA ZFAS1 在基因组水平上扩增(图 4),竞争性结合miR-150,抑制 ZEB1、MMP14 和 MMP16 基因表达以促进转移。另一种 ceRNA—FTX,除了结合 DNA 复制许可子 MCM2 外,还通过竞争性结合海绵状 miR-374a 和抑制 HCC 细胞 EMT 和侵袭发挥作用,从而阻碍 DNA 复制并抑制 HCC 细胞的增殖。 如上所述,LncRNAs 和miRNA竞争性结合的模型经常被使用。但是,在研究中应用该模型时,应当谨慎解释该模型的作用机制。这是因为该转录本身和其他问题导致的潜在的不平衡。尽管如此,PTAR,lncRNA-PNUT和lncRNA-ATB已被证明在多种癌症类型中发挥作用。
图4 转移相关的长链非编码 LncRNA 受各种上游机制的调控
转移定植的调节 一些 LncRNA 与促进肿瘤转移细胞在远处器官部位定殖和分化有关。
MALAT1与转移定植
MALAT1 是最早在癌症转移过程中被研究的 LncRNA 之一,它与肺癌转移有关,并且对早期非小细胞肺癌患者的生存率也有预测作用。在人管腔 B 型乳腺癌的小鼠乳腺肿瘤病毒 (MMTV)-多瘤病毒T 抗原 (PyMT) 模型中,启动子缺失或 ASO 介导的敲除MALAT1 基因可减少肺转移灶的形成,肿瘤分化更好(如囊性和包裹性肿瘤)。
此外,当 A549 肺癌细胞中的 MALAT1 表达受到抑制时(通过将前多腺苷酸化信号插入 MALAT1 基因座),各种转移相关基因表达降低(例如,ROBO1、MIA2、GPC6、LPHN2和 ABCA1),与对照细胞相比,体外转移减少。此外,小鼠尾静脉注射MALAT1 敲除肿瘤,肺转移减少,肿瘤细胞更多停留在血管内,表明它们无法侵入组织。研究结果提示MALAT1可作为靶点治疗转移性肺癌的潜力,皮下注射MALAT1 ASO可以降低转移性非小细胞肺癌肺小鼠模型中肺结节的肿瘤负荷。 尽管 MALAT1 通常被认为是一种促肿瘤转移的LncRNA,但一些证据表明 MALAT1 有助于抑制转移。在 MMTV-PyMT 小鼠模型中,插入早多聚腺苷酸序列使 MALAT1 的转录失活促进了肺转移,这与早期的实验结果形成鲜明对比。Kim 等人通过 MALAT1 的转基因过表达补偿转移效应后MALAT1 耗竭。在这项研究中,当使用现代技术(如质谱法全面鉴定 RNA 结合蛋白(ChIRP-MS )进行分析时,发现MALAT1可以与转移前TEAD转录物结合并使其失活。这可以导致转移和侵袭所必需的 ITGB4 和 VEGFA 表达的消除。这些相互独立的研究结果表明,LncRNA 的功能不仅取决于细胞环境(如比较乳腺癌与肺癌中的 MALAT1 功能),还取决于其主要的作用机制(如基因表达的调节和剪接,或转录因子失活 )。
MALAT1 也揭示了LncRNA 研究中的一个重大挑战。
器官特异趋向性调节
转移性细胞表现出器官特异性趋向性,因为各种器官环境需要对播散性肿瘤细胞的生存进行特定的适应。换句话说,特定的器官环境更适合播散性肿瘤细胞的生存。这一理论得到了促器官特异性转移性特定蛋白编码基因支持(例如骨转移中的结缔组织生长因子(CTGF)),使播散的肿瘤细胞能够克服组织微环境的需求。这种器官特异性转移的一个突出问题是乳腺癌,如何以非随机方式转移到远处器官(例如,骨骼、肺、中枢神经系统(CNS)、淋巴结和胃肠道)。高达 70% 的乳腺癌有骨转移,而 10-30% 转移至脑部。作为一种普遍存在的疾病,乳腺癌促进了几种特征良好的体内小鼠模型的建立,从而能够深入研究肿瘤发展中的这一关键过程。有研究显示某些 LncRNA与这一器官趋向性相关,其机制也适用于其他原发性恶性肿瘤中的 LncRNA的机制(图 5)。
骨转移
原发性乳腺癌的骨转移中,恶性肿瘤细胞旁分泌释放CTGF激活了破骨细胞YAP途径,以诱导分化和骨溶解,从而促进骨转移以及定植。使用高通量RNA (RNAi) 筛选显示,MCF-7乳腺癌细胞中YAP1激活需要LncRNA MAYA (图5)。敲除人类乳腺癌骨转移 (BoM-1833) 细胞中MAYA可消除CTGF分泌,从而减弱了癌细胞诱导的破骨细胞分化和再吸收。从机制上讲,MAYA与LLG2和甲基转移酶NSUN6形成分子复合物共同作用,使Hippo途径中MST1甲基化。甲基化MST1激活YAP1调节基因。与对照组相比,小鼠移植MAYA敲除的BoM-1833细胞后骨转移更少。此外,静脉内注射MAYA静默核酸 (LNAs) 可降低已接种乳腺癌细胞或A549肺癌细胞的小鼠的骨转移,这表明MAYA可作为骨转移治疗的精准靶向。
脑转移
与骨转移相反,乳腺癌脑转移通过利用不同的途径来实现器官特异性定植。原发性乳腺癌的脑转移细胞将L1细胞粘附分子 (L1CAM) 表达为共选择性血管内皮细胞,以达到脑转移性生长。Lnc-BM是一种LncRNA,与乳腺癌肺或骨转移相比,脑转移灶中表达显著提高 (图5)。Lnc- BM表达与患者生存率呈负相关,与中枢神经系统复发正相关。小鼠心内注射Lnc- BM (通过RNAi或CRISPR-Cas9) 剪切的乳腺癌细胞可减少脑转移,同时减少了癌细胞对脑内血管的共同选择。从机制上讲,Lnc-BM通过STAT1/3磷酸化激活ICAM1表达,从而允许肿瘤以类似于L1CAM的形式共同选择脑内皮细胞。此外,一旦细胞在脑内定植,Lnc-BM介导JAK2激活,通过CCL2募集巨噬细胞,调节利于脑转移的多种途径。
与X染色体失活不同,与骨,肝或肺转移相比,LncRNA XIST在小鼠的乳腺癌脑转移中被抑制。小鼠心内注射XIST沉默子的MCF-7和sk-br3乳腺癌细胞后,脑转移增加。XIST基因敲除的MMTV-PyMT自发乳腺癌转移小鼠模型中,通过诱导EMT和激活MET,脑转移增加。XIST敲除还可导致外泌体miR-503的释放,从而诱导M2小胶质细胞的极化,改变转移微环境。 由于LncRNAs的表达和功能通常是细胞类型特异性的 (除外 MALAT1 和 XIST),LncRNA 在癌症转移的器官特异性趋向性中发挥重要作用也许不足为奇。然而,目前尚不清楚在转移定植之前是否已经触发特异性 LncRNA 调控程序,以指导细胞分化途径。抑或是LncRNA 可通过微环境维持转移细胞的稳定。
图5 LncRNA与转移器官特异性趋向性
LncRNAs与转移微环境 转移细胞到达远处器官部位时,形成转移细胞并与TME相互作用。TME包括多种过程,如血管生成、抑制和/或共刺激-先天性和适应性免疫系统的选择,以及基质细胞的重组以促进转移性生长。转移细胞的生长需要建立维持支持性微环境,包括基质细胞、先天性和适应性免疫细胞。TME通过抑制免疫系统、诱导血管生成、旁分泌信号和转移前细胞外基质沉积促进癌症转移。LncRNAs在肿瘤和转移微环境中的作用如图所示(图6)。
微环境中NF-κB 相关 LncRNA 细胞质 LncRNA可直接抑制乳腺癌细胞内 NF-κB 复合物,当其下调时肿瘤转移潜力增加(图6a),NF-κB-相关LncRNA(NKILA)也通过STAT1-组蛋白乙酰化依赖机制调节T淋巴细胞的内在抗肿瘤活性(图4,6b)。在乳腺肿瘤浸润性 T 细胞中,NKILA 抑制 NF-κB 信号传导,活化-诱导T细胞死亡(图6b)。乳腺癌异种移植小鼠模型中,使用shRNA敲除NKILA的CD8+T细胞可增加其肺渗透,并且上调T细胞穿孔素和CD107a的表达,增强其抗肿瘤活性。Lnc-Tim3可诱导CD8+T细胞耗竭,促进p53和RelA转录靶点的激活。
NKILA对肿瘤细胞和免疫细胞中的NF-κB通路均有负性调节作用。CamK-A通过缺氧时诱导钙内流IκB的降解激活乳腺癌细胞中的NF-κB途径(图6b)。这导致IL6、IL8和VEGF(以及其他基因)的基因转录激活,最终促进患者体内巨噬细胞的募集和血管生成。
图6 LncRNA和肿瘤微环境
LncRNAs对免疫细胞的调节
肿瘤逃避免疫监视的调节性T细胞也受到LncRNA活性的影响。在CD4 T细胞中,Lnc- EGFR与细胞质EGFR结合,使其稳定于下游信号转导。进一步诱导FOXP3,向调节性 T 细胞系分化。与免疫抑制群体一致,当在HCC的异种移植模型中植入 lnc-EGFR 过表达T 细胞时,与对照相比,肿瘤生长增加。 在膀胱癌中,LNMAT1 在淋巴结阳性膀胱癌中过度表达,是膀胱癌患者总生存期的预后指标。裸鼠足垫注射ShRNA 介导的 LNMAT1 敲除膀胱癌细胞表现为淋巴结转移减少。然而,LNMAT1 的功能并不是肿瘤细胞固有的,LNMAT1 转导的膀胱癌细胞能够诱导巨噬细胞活化,这种作用可以被 CCL2 中和抗体消除(图 6c)。LNMAT1 通过与 CCL2 启动子形成 DNA-RNA 三链体激活 CCL2 的表达,并促进转录活性组蛋白修饰 H3K4 甲基沉积。此外,巨噬细胞依赖性上调 VEGF-C路径过表达LNMAT1可增加体外淋巴管生成。因此,LncRNAs可以对微环境产生有效的非细胞自主效应。 LncRNAs是免疫细胞分化的关键调节剂。LncRNAs对于神经、肺、心脏和胚胎组织的发育是必需的。由于所有这些细胞都有助于转移微环境或转移前生理状态,其他LncRNAs也将参与这些过程。
转移瘤中LncRNAs的翻译潜能
转移性肿瘤患者的生存率很低,对治疗的耐药性仍然是有效治疗的主要障碍。寻找一种组织和癌症特异性表达物作为肿瘤从原发性向转移性疾病进展的预后标志物是改善癌症治疗新的方法。
LncRNAs是一种新的诊断和预后工具
LncRNA PCA3是非-侵袭性前列腺癌的前期诊断标志物。基于对1008名患者的微阵列分析,LncRNA SChLAP1过表达是前列腺切除术后 10 年内前列腺癌转移的独立预后因素。在手术时将SChLAP1应用于患者的风险分层,有助于制定针对侵袭性肿瘤的前瞻性医疗策略。SChLAP1是前列腺癌临床治疗应用的关键基因分型。SChLAP1结合并拮抗SWI/SNF复合物,导致广泛SNF5基因组减少以及其靶基因的表达降低。因此,在SCID小鼠模型心内注射SChLAP1敲除22Rv1前列腺癌细胞后,这些细胞的转移率降低。然而,SChLAP1的这一机制也受到了质疑,研究表明,SChLAP1的过度表达未将SWI/SNF复合物从染色质中排除,这表明 SChLAP1 在前列腺癌转移中的作用机制与 SWI/SNF 无关。
转移性癌症患者的临床预后差异很大,尤其是不同组织学分型的转移癌患者。在转移性肾细胞癌(RCC)患者中,与低lncARSR水平的肿瘤相比,高lncARSR表达预示总生存期较短。lncARSR通过阻断miR-34和miR-44,下调AXL和MET表达,从而对舒尼替尼产生耐药性。lncARSR以外泌体为载体穿梭于细胞间,lncARSR能够以旁分泌方式促进耐药性的形成在肾细胞癌原位异种移植模型中静脉注射抗lncARSR的LNA ASOs可提高舒尼替尼治疗的敏感性。
RAMS11与结肠癌患者不良预后相关。RAMS11表达上调促进了对氟西鲁定 (FUDR,治疗转移性结肠癌的化疗药物) 的耐药性。RAMS11 CRISPR敲除细胞对FUDR敏感性增强。需要进一步的研究将RAMS11的预后和预测效用转化为临床应用。 目前生物标志物研究仍处于早期阶段。与没有肿瘤转移的患者相比,SChLAP1在肿瘤转移患者中表达升高,但许多具有癌症或组织特异性表达的LncRNA表达不足以高到可作为可靠的生物标志物。转录测序日益普及将促进lncRNA生物标志物的发现。由于疾病进展和对治疗反应的机制复杂,单一LncRNA可能不是临床上最有用的生物标志物,需要结合蛋白质编码基因评估潜在LncRNA 生物标志物。展望未来,无创检测 LncRNA 生物标志物的方法以减少活检需求也很重要。使用LncRNA 生物标志物对于无法反复进行肿瘤活检的患者进行连续监测尤为重要。总体而言,SChLAP1、lncARSR 和 RAMS11 作为LncRNA 在癌症治疗反应中有显著的作用,也有作为预测预后生物标志物的用途。
LncRNA靶向治疗靶点
LncRNA在转移肿瘤体内临床前模型中显示出作为癌症治疗靶点的可能。如在体内使用 ASO将lncRNA MAYA、MALAT1 和 lncARSR 各自作为靶点进行沉默,以减轻小鼠模型的转移。由于治疗会造成体内 LncRNA 转录物浓度进一步降低,在使用 ASO 以LncRNA作为靶点治疗时,确保最小的脱靶效应至关重要。除了 RNase H 介导降解成熟 RNA外,预测ASO 治疗活性时应考虑ASO通过早期终止转录破坏靶 RNA。某些 ASO也具有肝毒性,ASO 化学修饰可以减轻这种影响以及脱靶效应。 如何优化ASO的给药方式以增加针对 LncRNA 或蛋白质编码基因的 ASOs 的影响是目前实际的问题。 尽管存在这些挑战,但令人欣慰的是,在癌症小鼠模型以及临床试验中(如脊髓性肌萎缩症患者中的 SMN2 147 和难治性淋巴瘤中的和 SMAD7 在克罗恩病中的STAT3),ASOs已作为以蛋白质编码基因为靶点的治疗药物使用。
目前的局限性
许多研究表明LncRNAs在整个转移过程中发挥了作用,但目前仍存有知识空白。细胞转化和远处定植之间的侵袭-转移级联反应包括外渗、血源性扩散、内渗和微转移灶的形成,LncRNAs在这些反应中的作用没有充分明确。 此外,本综述中讨论的大多数 LncRNA 都具有增强转移潜能的作用。这些观察性研究结果是由于LncRNA 固有生物学属性或是研究方法的偏差造成的仍有待确定。 LncRNA潜在抑制转移的作用需要进一步研究。LncRNA 在跨癌症类型转移中的机制仍不明确。可能是基于LncRNA 的细胞特异性功能。 LncRNA 生物学领域的另一个难题是,看似不同的LncRNA 往往会集中作用于一些已确立的蛋白质编码途径上,而不是独立的调节转移。 几十年来对蛋白质编码基因的研究使发现并融合关键转移调节基因成为可能,而对许多 LncRNA 的研究相对处于起步阶段,并且还没类似的 LncRNA 发现。尽管蛋白质-RNA 有多种相互作用方式,多种 RNA 序列可与蛋白质编码基因相互作用并发挥相似的作用,研究认为RNA 需要与PRC2相互作用才能起效,综上所述,即使是从基因组不同位置产生的具有不同序列的大量 LncRNA 也可以有效产生类似的转移途径。
结论
转移是造成癌症致死的主要原因,LncRNA 不仅是癌症发生发展中重要的调节因子,而且是转移发病机制中的关键因素。近年来,随着越来越多的 LncRNA 被发现,LncRNA 在侵袭-转移级联、远处器官转移定植和肿瘤微环境中发挥显著作用。Lnc RNA 通过多种机制对癌症通路产生影响:与促转移或抗转移蛋白复合物一起发挥作用以及与其他 RNA 或 DNA 直接相互作用。然而,目前仍有许多悬而未决的问题,如LncRNA对于CTC、前转移灶形成和基质特异性细胞的作用等。未来的研究需要针对上述作用,并进一步研究LncRNAs参与肿瘤转移的机制。LncRNA已成为癌症治疗的新的靶点,它们在癌症进展中的不同作用为在临床抑制转移提供了新的机会。以LncRNAs为靶点的体内治疗技术的成熟,将极大地促进针对转移的LncRNA疗法的实现。
编译:陈婉南;翁梅琳 审校:张军;缪长虹
参考文献: Liu, S. J., Dang, H. X., Lim, D. A., Feng, F. Y., & Maher, C. A. (2021). Long noncoding RNAs in cancer metastasis.Nature reviews. Cancer, 21(7), 446–460. https://doi.org/10.1038/s41568-021-00353-1
声明:古麻今醉公众号为舒医汇旗下,古麻今醉公众号所发表内容之知识产权为舒医汇及主办方、原作者等相关权利人所有。未经许可,禁止进行转载、摘编、复制、裁切、录制等。经许可授权使用,亦须注明来源。欢迎转发、分享。
不感兴趣
看过了
取消

人点赞

人收藏

打赏
不感兴趣
看过了
取消
©2012-2023 北京华媒康讯信息技术股份有限公司 All Rights Reserved. 注册地址:北京 联系电话:010-82736610
广播电视节目制作经营许可证 —(京)字第 17437号 京海食药监械经营备20200522号
京ICP备12011723号 京ICP证150092号
 京公网安备 11010802020745号
工商备案公示信息
互联网药品信息服务资格证书((京)-非经营性-2020-0015)
京公网安备 11010802020745号
工商备案公示信息
互联网药品信息服务资格证书((京)-非经营性-2020-0015)






您已认证成功,可享专属会员优惠,买1年送3个月!
开通会员,资料、课程、直播、报告等海量内容免费看!

打赏金额
认可我就打赏我~
1元 5元 10元 20元 50元 其它
打赏作者
认可我就打赏我~
扫描二维码
立即打赏给Ta吧!
温馨提示:仅支持微信支付!
已收到您的咨询诉求 我们会尽快联系您



































 010-82736610
010-82736610
 股票代码: 872612
股票代码: 872612




