
体内稳态(葡萄糖和铁代谢)的调节需要考虑肠道微生物群在其与宿主的双向相互作用中的作用。
编译:微科盟蔚蓝,编辑:微科盟居居、江舜尧。
微科盟原创微文,欢迎转发转载,转载须注明来源《微生态》公众号。
导读
铁对地球上生命的出现和维持至关重要。几乎所有生物体都竞争或协作获取铁,表明了铁对于维持代谢稳态的关键生物和生理过程的重要性。在人类和其他哺乳动物中,肠道微生物群的细菌组成成分与宿主铁摄取的双向相互作用塑造了宿主和微生物群的代谢。细菌功能影响宿主对铁的吸收,而宿主体内铁的摄入、铁缺乏和铁过量则影响细菌的生物多样性、分类和功能,导致细菌毒力发生变化。宿主-微生物相互作用的结果影响铁的系统水平、不同组织中的铁储存和宿主葡萄糖代谢。在宿主和微生物群之间的界面处,宿主先天免疫系统和调节铁的循环可溶性因子(即铁调素、脂质运载蛋白2和乳铁蛋白)的变化与代谢疾病相关。事实上,肥胖相关代谢功能障碍和胰岛素抵抗的患者表现出铁稳态失调和肠道菌群变化。从进化的角度来看,对葡萄糖和铁这两种重要营养素的追求可能推动了人类向对人类生存和健康最有效的途径和基因的进化。
论文ID
原名:The role of iron in host-microbiota crosstalk and its effects on systemic glucose metabolism
译名:铁在宿主-微生物群互作中的作用及其对全身葡萄糖代谢的影响
期刊:Nature Reviews Endocrinology
IF:47.564
发表时间:2022.8
通讯作者:José Manuel Fernández-Real
通讯作者单位:西班牙赫罗纳生物医学研究所(IDIBGI)
DOI号:10.1038/s41574-022-00721-3
综述目录
1 前言
2 宿主铁稳态
2.1 自我平衡的机制
2.2 可溶性因子
2.3 细胞机制
3 细菌铁稳态
3.1 细菌铁获取
3.2 细菌铁储存
3.3 细菌铁稳态的调节
4 膳食铁对微生物群的影响
4.1 缺铁对宿主的影响
4.2 铁过载对宿主的影响
5 微生物群影响宿主铁稳态
6 宿主-微生物群代谢相互作用
6.1 影响葡萄糖代谢的代谢物
6.2 减弱毒力的相互作用
6.3 代谢相关性脂肪肝病
6.4 铁和脂肪组织
7 进化观点
8 结论
主要内容
1 前言
铁元素不仅是人体的重要组成部分,也是生命发展的关键因素。根据生命起源的理论,陆地岩石中的铁含量可能有助于地球表面水分的保持。此外,铁是许多生物前体分子非生物合成的关键。在地球上最古老的生物体中,铁是许多代谢相关氧化还原酶(如固氮酶、氢化酶以及参与硫酸盐还原和甲烷生成的酶)的主要辅因子。 第一批真核生物和真核多细胞生物似乎出现在大氧化事件之后,随后海洋中铁变得不可溶解且稀缺。在新元古代氧合事件发生时,真核生物多样性也显著增加,这与铁不溶性的恶化有关。自地球上生命起源以来,铁的获取对所有生物体都至关重要。此外,在持续不断变化的细菌种群中,集中于铁摄取的“社会”互动(包括竞争、协同和欺骗)已经形成。真核生物中多细胞生物的发展似乎发生于大氧合事件和新元古代氧合事件之间,使生物体整个生命周期内在一定程度上实现营养物质的循环利用。实际上人体中每天循环利用的铁大约是饮食中获得铁的25倍。重要的是,不需要铁的生命形式极为罕见,目前已知只有两种形式:细菌病原体Borrelia burgdorferi(莱姆病的病原体)和乳酸杆菌物种(共生细菌)。 铁一直是生物体内能量交换的代币,可能也是人类进化的推动力。鉴于其在DNA合成以及其他关键过程(例如氧气运输、细胞生长和分化、电子传递、线粒体生物合成和活动)中的作用,铁是细胞分裂所必需的。值得注意的是,铁代谢的改变常见于肥胖、糖耐量受损和2型糖尿病(T2DM)患者,这与组织铁积累的增加有关(主要在肝脏中)。此外,肥胖相关代谢功能障碍和胰岛素抵抗患者的铁稳态失调与肠道微生物群组成和功能的改变有关,这些变化在使用抗糖尿病药物(例如二甲双胍)治疗后会发生变化。 先前已有综述针对动物模型回顾了宿主和细菌间通过铁发生的多重相互作用。
本综述聚焦于人体研究以及肠道微生物组对宿主葡萄糖代谢的潜在影响(和双向关系)。在简要总结人体对铁的感知和摄取后,我们回顾了调节铁稳态的体液和细胞机制以及这些机制如何影响全身葡萄糖代谢。我们探讨了外源性铁如何影响肠道微生物群的组成,以及肠道微生物群如何调节铁稳态。最后,我们讨论了宿主-微生物-铁代谢相互作用如何影响肠道和全身葡萄糖代谢、脂肪组织功能和肝脂肪变性的发展。
2 宿主铁稳态
2.1 自我平衡的机制
就其与微生物群和宿主免疫系统的相互作用而言,铁是最具特征的微量营养素之一。铁是人体中含量最丰富的金属之一,在血红蛋白和各种代谢酶(如铁硫簇)中作为蛋白质辅因子。经饮食摄入后,铁到达消化道,主要吸收场所为十二指肠(图1)。以三价铁形态(Fe3+)到达肠腔后,被肠上皮细胞顶膜的十二指肠细胞色素b(DCYTB)还原为亚铁(Fe2+)并被吸收。随后,Fe2+通过顶膜的二价金属转运蛋白1(DMT1)进入肠细胞。血红素载体蛋白1(HCP1)负责将血红素铁转运进入十二指肠肠上皮细胞,尽管HCP1在叶酸转运中的作用可能比血红素更重要。铁贮量可以调节HCP1蛋白的亚细胞定位而不影响其表达,而缺氧可显著上调编码HCP1(SLC46A1)的基因。血红素吸收后,血红素加氧酶1(HO1)降解血红素释放Fe2+。在肠细胞内部,铁可以留在细胞内或离开细胞进入血液。铁从肠上皮细胞的基底外侧膜通过膜铁转运蛋白1出胞,同时被亚铁氧化酶氧化生成Fe3+。
图1 参与宿主铁稳态的通路。
肠道血红素和非血红素铁吸收的主要机制。十二指肠是膳食铁吸收的主要部位,在肠腔中,三价铁(Fe3+)被十二指肠细胞色素b(DCYTB)还原为亚铁(Fe2+)。然后二价金属转运蛋白1(DMT1)将Fe2+转运到肠上皮细胞中。血红素铁也可通过血红素载体蛋白1(HCP1)进入十二指肠上皮细胞。随后血红素加氧酶1(HO1)从血红素中释放出Fe2+。在细胞内水平,铁与铁调节蛋白1(IRP1)和IRP2以及缺氧诱导因子2α(HIF2α)相互作用。HIF2α信号传导对DMT1、DCYTB以及铁转运蛋白的调节至关重要,后者将亚铁转运穿过肠细胞基底外侧膜。HIF2α受转录后调控,它与DMT1和DCYTB启动子调节区域中的缺氧反应元件(HRE)结合,并且包含一个铁反应元件(IRE)结构域。IRP1和IRP2与含IRE的mRNA(如FTH1、FTL、FPN、TFR1、DMT1和HIF2A)结合抑制这些转录翻译。细胞内的铁储存由铁蛋白控制,而Fe2+通过铁转运蛋白输出到血液中,同时被亚铁氧化酶氧化成Fe3+。循环中的Fe2+也被铜蓝蛋白氧化成Fe3+。血浆转铁蛋白捕获Fe3+并将其分配到全身所有细胞(holo-transferrin)。细胞通过TFR1介导的holo-transferrin内吞作用摄取铁。
在红系前体细胞中,DMT1将铁转运到细胞质中,然后穿梭到线粒体中进行血红素生物合成。脾脏网状内皮巨噬细胞可能通过TFR1或回收衰老红细胞而获得铁。全身铁稳态由肝铁调素(由HAMP编码)的产生调节。铁调素结合并诱导铁转运蛋白的降解。 血液中的三价铁迅速与糖蛋白转铁蛋白结合,转铁蛋白是循环铁的主要形式。转铁蛋白将铁分配给体内所有外周细胞以满足其需要。转铁蛋白受体1(TFR1)存在于所有细胞中,吸收Fe3+-转铁蛋白复合物。进入任何细胞后铁就会储存在铁蛋白中(铁代谢的核心蛋白质)。铁被吸收并被上皮细胞中的铁蛋白捕获后,铁被排除在身体其余部分之外,并通过成熟肠细胞的脱落逐渐消除。铁蛋白主要是一种铁储存蛋白,通过其信使RNA中的铁反应元件(IREs)受细胞铁状态的转录后调控(图1)。较高的细胞内铁浓度会导致铁蛋白表达增加,而铁缺乏时较低浓度抑制其表达。但铁蛋白也是一种急性期蛋白,在炎症条件下血清浓度增加。肝损伤后铁蛋白从肝细胞中渗出,导致血浆浓度升高。在稳态条件下血清中可测量的铁蛋白似乎主要来自巨噬细胞,反映了铁的全身储存;即铁蛋白的血浆浓度反映了肝脏和其他组织中的浓度。
肝脏分泌铁调素(全身铁代谢最重要的循环调节剂之一),它通过诱导网状内皮巨噬细胞和肠上皮细胞表面的铁转运蛋白降解而发挥作用。铁超负荷刺激铁调素的表达,从而抑制铁输出到循环中并阻止衰老红细胞中铁的回收。细胞内铁稳态也受十二指肠缺氧诱导因子2α(HIF2)及铁调节蛋白1(IRP1)和IRP2的调节。细胞内缺铁条件下HIF2α转录上调肠上皮细胞DCYTB、DMT1和FPN的表达,而IRP与IREs结合并在转录后调节许多铁稳态基因的表达,包括HIF2A。从机制上讲,IREs是位于铁蛋白mRNA的5'-非翻译区(UTR)和转铁蛋白受体mRNA 3'-UTR中的茎环结构。IRP与IREs结合会破坏铁蛋白mRNA的翻译,而转铁蛋白受体mRNA的稳定性会增加,从而增加细胞膜中转铁蛋白受体的水平和细胞对铁的摄取。在肠细胞中,IRPs负调控铁蛋白翻译以防止铁螯合。值得注意的是,铁在免疫系统中具有重要作用(Box 1)。
Box1 铁在免疫系统中的作用。
在宿主防御中,铁参与活性氧和氮中间体的产生,并作为先天免疫和适应性免疫的调节剂。在先天免疫反应中,呼吸爆发需要铁,这种呼吸爆发可以杀死吞噬细胞内的病原体,当铁受到限制时,吞噬细胞会受损。小鼠铁缺乏还与T细胞应答的下调有关,例如抗原特异性反应和多克隆增殖。此外,铁缺乏会导致T细胞上CD28的表达减少,这是一种在T细胞活化中起重要作用的共刺激受体。事实上,缺铁小鼠对实验性自身免疫性脑脊髓炎的发展具有抗性,迟发性超敏反应降低,血清IgM和IgG水平降低。一些小鼠模型已经证明铁在抗感染中的重要性。Nramp1突变小鼠更容易感染Salmonella typhimurium、Mycobacterium bovis Bacillus Calmette–Guerin和Leishmania major。另一方面,分枝杆菌、Chlamydia species和Legionella pneumophila的细胞内生长主要依赖于宿主来源的铁。为了应对损伤,宿主激活体液和细胞机制,降低铁的可用性,进而减弱病原体的增殖。
2.2 可溶性因子
20世纪40年代就已观察到在培养基中添加生蛋清会严重抑制多种细菌和真菌的生长。在蛋清存在的情况下,单独补充铁就足以恢复微生物的生长。蛋清中存在的铁结合成分似乎与微生物争夺铁。两年后在人血清中也发现了类似的抗菌活性。介导生鸡蛋和血液中这种活性的因素是转铁蛋白。铁蛋白也被证明与秀丽隐杆线虫(Caenorhabditis elegans)的先天免疫有关。除了转铁蛋白和铁蛋白外,其他可溶性蛋白质也在不断地感知铁的可用性并调节体内铁稳态。与肥胖状态、全身葡萄糖代谢、胰岛素抵抗和T2DM相关的研究中最多的是铁调素、脂质运载蛋白2和乳铁蛋白。
2.2.1 铁调素
膳食铁过量及来自革兰氏阴性菌的全身性脂多糖(LPS)诱导铁调素的表达。铁调素对循环铁和组织铁可用性的精准调控与预防细菌感染有关。铁调素基因的缺失导致小鼠铁过载,而转基因过表达可导致贫血。铁调素下调几种细胞类型和组织(例如巨噬细胞、小肠、肝脏和脂肪组织)中的铁转运蛋白mRNA和蛋白质水平。铁调素也受骨形态发生蛋白6和瘦素的调节。此外,肥胖个体的铁调素水平增加,同时肝脏铁含量和铁积累的循环标志物(血清铁蛋白水平)也增加。内脏脂肪组织与总脂肪组织的比例与铁调素水平的增加密切相关,同时铁吸收和利用率降低。在铁代谢参数中这些关联表现出普遍的性别二态性,众所周知与女性相比男性的铁储存量增加。严重肥胖(BMI>35 kg/m2)和新诊断的T2DM个体的血清铁调素水平与小肠中铁转运蛋白的表达呈负相关,这些个体中参与非血红素铁吸收的基因表达增加。 相比之下,T2DM患者的循环铁调素减少。通过在大鼠中使用链脲霉素诱导糖尿病导致血清铁调素水平降低、肠道中铁转运蛋白表达增加以及铁吸收增加,从而导致血清和肝脏铁含量增加。
2.2.2 脂质运载蛋白2
上皮细胞和中性粒细胞分泌脂质运载蛋白2,它通过一组称为铁载体的小分子铁螯合剂与铁复合。与这些分子相互作用后,铁载体-脂质运载蛋白2复合物被内化并转运到晚期胞内体,促进细胞和组织对铁的吸收。这种机制构成了在发育过程中活跃的铁传递途径(如发育上皮细胞中),这在细胞生理学中很重要,并且独立于转铁蛋白-内吞途径。 当铁含量受限时,这种脂质运载蛋白2途径变得至关重要。许多微生物通过铁载体的合成、分泌和再摄取从环境中获取铁,铁载体与铁结合的亲和力远高于内源性载体蛋白转铁蛋白,从而有利于感染期间病原体的存活。在细菌与宿主间的军备竞赛中,内源性蛋白脂质运载蛋白2通过螯合铁发挥强大的抑菌作用。由于细菌同时利用细菌和哺乳动物的铁载体来输入铁,宿主通过抑制铁载体合成和通过toll样受体信号传导诱导脂质运载蛋白2的表达来应对感染。
人体中 脂质运载蛋白2的循环浓度与肥胖、高甘油三酯血症、高血糖、胰岛素抵抗和C反应蛋白的循环水平正相关,且与年龄和性别无关。T2DM患者使用罗格列酮治疗后血浆脂质运载蛋白2浓度降低,这与胰岛素敏感性增加(P=0.002)和血清C反应蛋白浓度降低显著相关(P=0.003)。在另一项独立研究中,脂质运载蛋白2的血清浓度也与空腹甘油三酯显著相关。膳食脂肪过量后,循环脂质运载蛋白2水平也急剧增加,这与严重肥胖个体的空腹胰岛素(P<0.001)、HOMA-IR (P=0.02)和餐后甘油三酯浓度 (P=0.018)显著相关。
2.2.3 乳铁蛋白
乳铁蛋白是一种存在于体液(如乳液、唾液、泪液和血清)中的高亲和力铁结合蛋白,由中性粒细胞在炎症部位脱颗粒时释放出来,通过螯合铁来抑制细菌生长。与铁调素和脂质运载蛋白2相比,乳铁蛋白似乎与人体铁稳态的调节无关。然而乳铁蛋白可能通过间接机制对全身代谢产生有益影响。 细菌病原体变形链球菌(Streptococcus mutans)和单核细胞增多性李斯特菌(Listeria monocytogenes)表达膜结合或分泌的铁还原酶,其作用是从乳铁蛋白和转铁蛋白中释放铁。给肥胖小鼠服用乳铁蛋白可降低肝脏铁含量,同时改善血脂异常、降低胰岛素抵抗、改善葡萄糖耐量和肝脏脂肪变性。除了人体观察性研究,在小鼠和大鼠中的其他实验研究也支持乳铁蛋白在代谢疾病中的这种保护作用。在人体研究中,乳铁蛋白的循环浓度与空腹甘油三酯浓度、腰臀比和空腹葡萄糖浓度呈负相关,并与HDL水平直接相关。在另一项针对有或无糖耐量改变个体的研究中,循环乳铁蛋白与胰岛素敏感性呈正相关,与炎症标志物呈负相关。与健康个体相比,T2DM个体中LPS诱导的中性粒细胞乳铁蛋白释放显著降低,而罗格列酮体外处理全血导致中性粒细胞分泌乳铁蛋白增加。
2.2.4 总结
这些研究表明循环铁调素、脂质运载蛋白2和乳铁蛋白可能部分解释了在肥胖、糖耐量受损和/或T2DM个体中观察到的铁稳态失调,这导致肝脏、脂肪组织和骨骼肌中组织铁积累增加。长期铁过量诱导的氧化应激可能会损害组织胰岛素作用并促进全身胰岛素抵抗和高血糖。在恶性循环中,铁调素和乳铁蛋白的循环水平降低会进一步加重,这种作用可能会导致肠道铁吸收增加和脂质运载蛋白2相关有害作用。事实上,在T2DM中已观察到与微血管和大血管并发症相关的脂质运载蛋白2循环浓度的增加。
2.3 细胞机制
在细胞水平上,巨噬细胞在全身铁稳态中发挥关键作用,其为红细胞生成提供铁或螯合铁用于细菌抑制。本综述总结了巨噬细胞和铁稳态在代谢中的相关性。 当小鼠巨噬细胞通过铁转运蛋白的特异性骨髓靶向缺失抑制铁输出时,组织铁在骨髓、肝脏和脂肪组织中积聚。在老年小鼠中,铁积累导致肝脏和脂肪组织巨噬细胞线粒体呼吸功能受损,进而导致葡萄糖耐量受损。在骨髓特异性铁转运蛋白缺失小鼠的脂肪组织中,巨噬细胞被重新配置并获得较高的铁回收能力,保护脂肪细胞免受铁过量和炎症的有害影响。值得注意的是,小鼠的肥胖与铁再循环能力低的巨噬细胞增加和铁处理基因表达减少有关,同时脂肪细胞铁储存增加四倍;肝脏铁储备也随之减少。小鼠巨噬细胞铁转运蛋白的选择性破坏也导致了肌肉浸润性巨噬细胞内铁的积累,导致肌肉中更小的肌纤维和脂肪组织的积累。 小鼠巨噬细胞中编码铁蛋白重链1 (Fth1)基因的特异性缺失导致细胞内铁浓度降低,且由于铁输出增加而减少脂肪组织和脾脏中的铁积累。这种表型与体重减轻、脂肪组织炎症和氧化应激有关。高脂饮食后,与野生型小鼠相比,巨噬细胞Fth1缺失的小鼠胰岛素敏感性也有所提高。
3 细菌铁稳态
铁是人体的重要营养素,也是许多生态系统中限制生长的营养素。膳食铁仅5-20%被十二指肠吸收,因此摄入的铁有大约80%留在肠腔中(主要是结肠),供微生物群利用。肠道微生物群中几乎所有细菌的复制和生长都依赖于对这种未吸收膳食铁的竞争,因为铁是参与关键代谢途径(例如氧化还原反应、电子传递链、DNA合成和短链脂肪酸(SCFAs)的产生)的蛋白质的辅因子。铁对多种病原体的毒力和定殖也有重要影响。因此,细菌蛋白质组中多达25%的蛋白质使用铁作为辅因子来实现其细胞功能。然而,涉及的相同氧化还原过程通过产生对生物分子(脂质、蛋白质和DNA)造成损害的活性氧而使铁具有潜在毒性。 鉴于游离铁具有很强的毒性和活性,它与高亲和力宿主蛋白如转铁蛋白、铁蛋白、乳铁蛋白或血红素蛋白结合。因此,体液和肠腔中的游离铁浓度低至~10-24 M,远低于细菌最佳生长所需的10-6 M浓度。在肠道的非酸性pH值下,游离铁在很大程度上是不溶的。在这些铁的限制条件下,微生物与宿主之间以及微生物物种之间对铁的争夺非常激烈。因此,需要严格调控细菌铁稳态以维持足够的不稳定铁储备,同时避免过量的有毒铁以防止细胞损伤。
3.1 细菌铁获取
为了克服对铁的竞争压力,微生物已经进化出几种复杂的策略来获取铁(图2)。几乎所有已知的细菌种类都能产生铁载体。典型的细菌铁载体是肠杆菌素儿茶酚酸盐,它参与宿主转铁蛋白中铁的回收。然而,哺乳动物会产生铁载体样代谢物和隔离肠杆菌素的脂质运载蛋白2。因此,微生物具有多种铁载体系统,已经描述了数百种铁载体。重要的是,铁载体可以在微生物群落中产生巨大的适应性效果。一种微生物分泌的铁载体可能会在缺乏合适受体的物种中引起铁饥饿,但能令具有匹配受体(异铁载体)的其他微生物获得铁。
图 2 革兰氏阴性菌的铁稳态。
细菌已经进化出几种获取铁的机制,其中最普遍的是铁载体的合成和分泌。这些因子是高亲和力的Fe3+清除小分子螯合剂。孔蛋白在能量转导TonB-ExbB-ExbD系统的介导下,将Fe3+-铁载体复合物转运穿过细菌外膜。微生物有两种获取血红素的机制:直接摄取游离血红素或与宿主蛋白质(血红蛋白或肌红蛋白)结合的血红素,并分泌结合血红素的haemophores。haemophores将提取的血红素输送到外膜,也以TonB-ExbB-ExbD依赖的方式运输。或者,许多细菌病原体具有特定的外膜受体以利用宿主非血红素、含铁蛋白质,例如转铁蛋白(如TbpA或TbpB)或乳铁蛋白(如LbpA或LbpB)。进入外周胞质后周质结合蛋白(PBP)将铁载体、血红素或三价铁转运到细胞质膜,在此它们通过ABC渗透酶(例如FepCDG、HemUV或FbpBC)转移到细胞质中。细菌已经进化出直接获取亚铁(Fe2+)的机制(主要在厌氧或酸性条件下),例如Feo系统。当不稳定铁库超过细胞需求时,可以将其储存起来,避免活性氧形成。储存蛋白(来自饥饿细胞的铁蛋白、细菌铁蛋白或DNA结合蛋白)形成一个球形蛋白外壳,包围着储存铁的中央空腔。细菌铁稳态的严格调控主要通过铁摄取调节因子(Fur)家族成员实现。在铁充足的情况下,Fur通过将Fur-Fe2+复合物结合到称为“Fur box”的调节区域来抑制铁获取基因的转录。大肠杆菌的代表性铁吸收系统显示在括号中。HO1:血红素加氧酶 1。
3.2 细菌铁储存
与获取机制无关,铁主要以Fe2+的形式进入不稳定铁库。不稳定铁库是铁依赖性蛋白质组的正常功能所必需的,但当它超过代谢需求时,由于活性氧的形成,游离铁会产生毁灭性的后果。因此,细菌具有三种属于铁蛋白超家族的专用铁储存蛋白:细菌铁蛋白,类似于真核铁蛋白;含血红素的铁蛋白,称为细菌铁蛋白,仅存在于细菌中;以及来自饥饿细胞的更小的DNA结合蛋白,也称为微型铁蛋白,仅存在于原核生物中。
3.3 细菌铁稳态的调节
对细菌铁稳态的严格调控对于确保足够的铁平衡以满足代谢需求同时抑制由于活性氧的产生引起的细胞损伤至关重要。铁摄取调节因子(Fur)是一种Fe2+结合金属蛋白,是控制细菌中铁稳态的最广泛、保守的主要转录调节因子(图2)。在细菌胞内不稳定铁库达到可接受的水平后,Fur会抑制铁获取基因的转录。Fur也被证明可以直接和间接地激活基因表达。值得注意的是,高鸟嘌呤-胞嘧啶含量的革兰氏阳性细菌使用白喉毒素调节蛋白来控制铁依赖性基因的表达,而α-变形菌中的铁调控主要由两种转录调节因子(铁反应调节因子和根瘤菌铁调节因子A)完成。
4 膳食铁对微生物群的影响 对铁的持续争夺有可能彻底改变肠道细菌生态系统。
例如,缺失调控小鼠全身铁稳态的关键基因(Irp2、Hfe)会导致肠道微生物群组成发生改变。与Hfe-/-小鼠相比,Irp2-/-小鼠的粪便铁含量增加,Lactobacillus murinus和Lactobacillus intestinalis的相对丰度更高,而Hfe-/-小鼠中屎肠球菌(Enterococcus faecium)高度占优势。此外,在Hfe-/-小鼠中观察到结肠微生物群的显著变化。Hfe-/-中来自变形菌门和TM7门的病原菌相对丰度高于野生型小鼠,其中来自普雷沃氏菌属(Prevotella)、肠球菌属(Enterococcus)、链球菌属(Streptococcus)、Turicibacter、Sutterella和脱硫弧菌属(Desulfovibrio)的“促炎”细菌增加。孕妇中铁状态的生化标志物,如血清铁蛋白水平和转铁蛋白饱和度,与共生双歧杆菌科水平降低和致病性肠杆菌科和大肠杆菌增加相关。
4.1 缺铁对宿主的影响
有益的共生细菌(乳酸杆菌和双歧杆菌)对铁的需求很少或根本不需要,它们中的大多数不产生铁载体或任何其他铁载体。在小鼠和人类微生物群中这些细菌的富集与肠道屏障功能改善和肠道病原体定植的减少有关。与这一发现一致,猪在生命早期缺铁导致乳酸杆菌和双歧杆菌的相对丰度增加。饮食铁缺乏后,小鼠(十二指肠和粪便)中约氏乳杆菌(Lactobacillus johnsonii)和罗伊氏乳杆菌(Lactobacillus reuteri)显著增加,而在大鼠体内观察到乳杆菌科和肠杆菌科增加以及Roseburia和Eubacterium rectale(丁酸生产菌)大量减少。野生型和杂合TNFΔARE/WT小鼠(一种克罗恩病模型)中肠腔铁的消耗导致双歧杆菌属、Succinivibrio、Turicibacter和梭菌属的数量增加,且还在野生型和结肠炎易感小鼠中诱导肠杆菌科细菌(例如大肠杆菌)的扩增。 体外实验进一步支持了动物模型的结果;例如,结肠发酵研究表明,在铁螯合或极低铁浓度的培养基中,双歧杆菌科、乳杆菌科以及肠杆菌科的数量显著增加。与共生微生物相比,低浓度铁条件下致病性肠杆菌科的持续增加可能源于更有效的铁获取能力。因此来自肠杆菌科的病原体(如大肠杆菌、沙门氏菌Salmonella和肺炎克雷伯菌Klebsiella pneumoniae)会分泌改良形式的铁载体肠杆菌素,它可以逃避宿主脂质运载蛋白2在啮齿动物炎症肠道中的隔离。因此,分泌改良肠杆菌素的病原体的生长优于仅依赖肠杆菌素的铁载体介导铁获取的共生菌。
4.2 铁过载对宿主的影响
乳酸杆菌和双歧杆菌在铁含量有限的环境中生长繁殖,但在富含铁的环境中被其他微生物取代。
Ivorian学校儿童(6-14岁)的铁强化导致肠道菌群发生变化,肠道炎症发生的同时乳杆菌科微生物减少,肠杆菌科微生物增加。在6个月大的肯尼亚婴儿中观察到类似的效果:补充含铁粉有利于大肠杆菌和志贺氏菌属(Shigella)(肠杆菌科)的生长,增加肠杆菌与乳杆菌的比例,加剧肠道炎症,并降低双歧杆菌的相对丰度。在西方人群(9-10个月大的美国婴儿)中,与含铁较少的肉类饮食相比,含铁强化谷物的饮食降低了双歧杆菌属、乳杆菌目和罗氏菌属(Rothia)的丰度。 与对照饮食的小鼠相比,补充膳食血红素后肠杆菌科微生物丰度更高。在结肠炎小鼠模型中,高水平膳食铁导致变形菌门细菌数量显著增加并加剧炎症。在大鼠中,口服铁剂(灌胃1 mL赋形剂(0.01 mol/L HCl)或1 mL含8 mg、16 mg或24 mg铁的液体铁制剂30天)可增加肠道菌群中共生瘤胃球菌科、Defluvitalaceae和粪球菌属(Coprococcus),减少毛螺菌科。 过量铁的影响还取决于背景饮食。例如,我们发现12周龄雄性C57BL/6小鼠肠道微生物群的细菌多样性和观察到的物种数量随着饮食中脂肪和铁含量发生显著变化。
正如预期,高脂肪饮食会降低细菌生物多样性,但仅在低铁饮食中出现。高脂肪和高铁含量饮食会导致细菌生物多样性增加。值得注意的是,当饮食中铁含量增加时,高脂肪饮食和对照饮食之间肠道细菌组成的差异低于饮食铁含量低时,且在高铁水平下变得不显著。 血红素摄入是功能性铁吸收的重要途径,占人体铁摄入量的三分之二。流行病学研究表明高血红素摄入会增加T2DM的患病风险,并且与体内储存的铁量呈正相关。从机制上讲,过量的细胞内游离血红素会催化活性氧的形成并促进氧化应激,从而诱导β细胞凋亡、肝功能障碍和胰岛素抵抗。小鼠研究表明,膳食中添加血红素会导致相关的肠道微生物群失调,其特征是细菌α多样性(衡量生态系统内物种多样性的指标)降低、厚壁菌门减少以及变形菌门丰度增加,尤其是肠杆菌科。这种微生物群失调与肠道炎症增强有关。 在体外模型中,在人TNO肠道模型2(一种大肠的体外模型)中补充铁也诱导微生物群组成的转变,降低了乳杆菌、双歧杆菌以及其他类型乳酸菌(如链球菌Streptococcus、片球菌Pediococcus和肠球菌Enterococcus)的水平。
5 微生物群影响宿主铁稳态
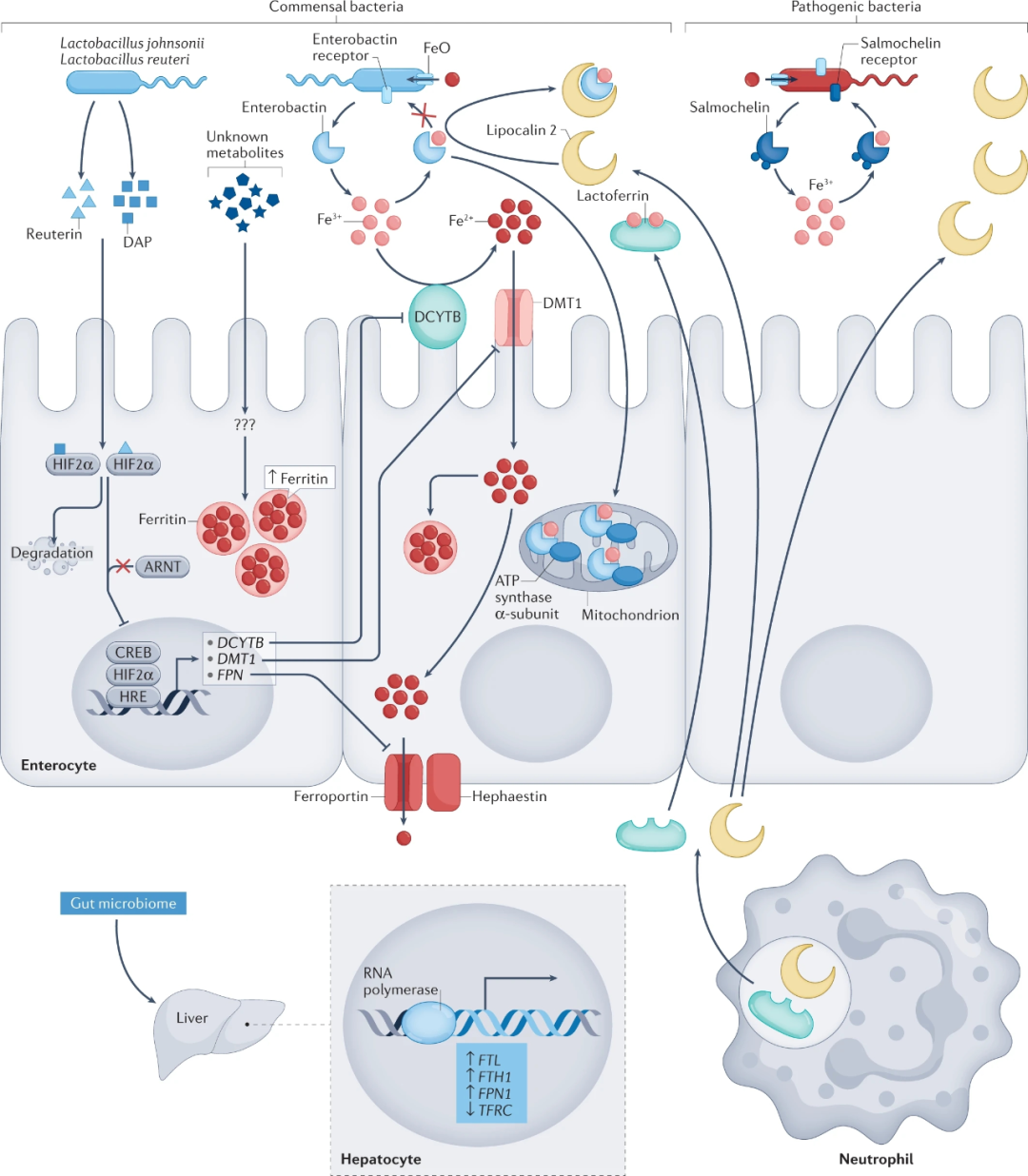
肠道微生物群本身似乎就在铁稳态的调节中发挥作用(图3)。
早期研究表明,无菌但非常规饲养的大鼠在喂食低铁饮食时会出现贫血。与常规饲养的大鼠相比,无菌大鼠的铁吸收减少,导致粪便中铁的流失增加。无菌小鼠的微生物群定植导致参与顶端铁摄取的蛋白质(DCYTB和DMT1)的表达降低,同时铁蛋白和铁转运蛋白表达增加,细胞铁储存和输出增加。2020年的一项研究证实了类似结果,该研究还发现无特定病原体小鼠中抗生素治疗会导致肠道铁转运蛋白DCYTB和DMT1的表达水平升高,但铁蛋白表达水平降低,这进一步证实了肠道微生物群在铁稳态中的作用。从机制上讲,原生微生物群通过抑制HIF2α(肠道铁吸收的主要转录因子)功能,以不依赖于铁调素的方式抑制肠道铁转运蛋白的表达。然而,肠道微生物群对铁蛋白的调控与HIF2α无关。共生微生物产生的铁载体不仅增加了细菌的铁吸收,而且影响了宿主的铁稳态。例如肠杆菌素通过与宿主线粒体ATP合酶的α亚基结合来提升宿主线粒体铁水平,从而抵消肠杆菌素的铁清除作用并促进宿主发育。在结肠炎小鼠模型肠道炎症的背景下,共生微生物群还调节肝脏铁调素的表达。 从人类到小鼠的粪便微生物群移植表明,铁代谢也可以通过肠道微生物群表型进行复制。从具有高-正常血清铁蛋白水平个体移植微生物群,导致受体小鼠肝脏中与铁代谢相关的基因发生改变,Ftl1、Fth1和Slc40a1的表达增加,转铁蛋白受体(Tfrc)表达降低。
Caco-2细胞(一种人类结直肠腺癌上皮细胞系)的体外研究表明,微生物组通过产生有机酸(如SCFAs和乳酸)从而降低结肠的pH值并有利于更易溶解的亚铁的存在,进而增强结肠铁的吸收。例如,补充植物乳杆菌Lactobacillus plantarum 299v菌株可显著增加育龄期女性从果汁饮料中吸收的铁增加约50%。SCFAs还可调节上皮细胞中铁代谢基因的表达,刺激上皮细胞增殖。微生物群介导的鞣花酸向尿石素A的转化也被证明可以增强小鼠对铁的吸收。
图3 肠道菌群对宿主全身铁稳态的调节。
肠道缺铁情况下微生物群中的共生细菌(如乳酸杆菌)会分泌代谢物(如1,3-二氨基丙烷(DAP)),通过破坏HIF2α-ARNT来抑制缺氧诱导因子2α(HIF2α),从而导致HIF2α降解。随后,肠道转运蛋白DCYTB、DMT1和铁转运蛋白转录下调。同时其他未鉴定的微生物代谢物通过未知机制上调铁蛋白的表达。许多共生和致病细菌已经进化出复杂的机制从宿主中获取铁。细菌可以通过FeO系统直接吸收亚铁(Fe2+)。或者它们产生铁螯合铁载体(例如肠杆菌素),以获取三价铁(Fe3+)。作为响应,宿主的肠细胞和免疫细胞产生铁螯合蛋白(例如乳铁蛋白)与Fe3+结合。宿主细胞还分泌脂质运载蛋白2,它可以隔离肠杆菌素。一些来自肠杆菌科的病原菌也可以释放一种称为沙门菌素的改良肠杆菌素,它不受脂质运载蛋白2的抑制。肠杆菌素的清除作用可能会对宿主体内的铁稳态和细胞过程产生负面影响。然而由共生细菌产生的肠杆菌素被证明通过与宿主线粒体ATP合酶α亚基相互作用以协同机制促进宿主线粒体铁的获取,从而增加线粒体铁水平以及低铁和高铁环境中的宿主发育。研究表明,高浓度血清铁蛋白的人类供体粪便菌群移植可改变受体小鼠肝脏中铁相关基因(ftl1、fth1、fpn1和tfrc)的表达。
6 宿主-微生物群代谢相互作用
肥胖相关代谢紊乱和胰岛素抵抗的患者表现为铁稳态失调和肠道菌群失调;因此,在代谢疾病的发展过程中铁稳态的改变与肠道微生物组之间可能存在联系(图4)。由铁引起的微生物组组成变化可能会导致实质性的功能改变,这可能发生在本节总结的以下三个层面。
图4 肠道微生物群、铁和葡萄糖代谢之间的相互作用。
在小鼠感染鼠柠檬酸杆菌Citrobacter rodentium(啮齿动物粘膜病原体)期间口服补充羰基铁可诱导全身胰岛素抵抗,这与白色脂肪组织中的铁过载有关。这种胰岛素抵抗降低了肠道内葡萄糖的吸收并增加了肠腔中葡萄糖浓度。这一效应增加了肠道中病原体对葡萄糖的利用并抑制了毒力因子的表达,从而筛选出了减毒C. rodentium菌株。显然,与死亡的感染小鼠或未感染的对照小鼠相比,感染后存活小鼠的肝脏转录组富含铁代谢基因。疟疾和一些细菌感染导致广泛溶血;随后细胞外血红蛋白被氧化并释放血红素。血红素的血浆积累会破坏免疫细胞功能,并可能导致器官损伤,最终导致败血症。这种效应被血红素加氧酶1(HO1)的诱导作用所抵消,HO1催化血红素的分解并释放铁。不稳定的铁会导致肝细胞氧化应激并抑制葡萄糖-6-磷酸酶(G6Pase),从而导致致命的低血糖。铁蛋白通过肝糖异生对抗G6Pase抑制并维持血液中的葡萄糖水平,防止低血糖并建立感染耐受性。在人类中,血清铁蛋白水平与代谢相关性脂肪肝疾病(MAFLD)的晚期阶段、糖异生代谢物以及厚壁菌门、放线菌门和变形菌门科的富集密切相关,特别是Micrococcaceae、Leuconostocaceae和Pasteurellaceae。MAFLD分期、这些细菌科和血清铁蛋白水平同时与包括GYS2、SEC24B、SLC51A和ACSM5在内的特定肝脏特征相关。尤其是GYS2(催化糖原合成中的限速步骤)与晚期MAFLD具有最强的负相关性。SEC24B已被证明有助于内质网应激、肝损伤和MAFLD。SLC51A参与胆汁酸(主要来源于微生物)运输,这也与MAFLD相关。与具有更健康的微生物群组成的对照小鼠相比,敲除小鼠脂肪细胞中的Tfrc可降低脂肪细胞中的铁含量并诱导更健康(更少炎症)的白色脂肪组织。ROS,活性氧。
6.1 影响葡萄糖代谢的代谢物
微生物组具有广泛的代谢活性并产生广泛的生物活性代谢物,这些代谢物来源于碳水化合物(SCFAs和琥珀酸盐)和蛋白质发酵(支链脂肪酸、酚类和吲哚)、甲胺、胆汁酸、有机酸和维生素。因此,微生物群与宿主之间的相互作用主要由这些微生物衍生的代谢物介导,这些代谢物通过影响体重、能量稳态、脂质和葡萄糖代谢以及炎症状态,在微生物群和宿主健康间发挥中介作用。 微生物衍生代谢物在全身代谢中的作用已在别处讨论过。在葡萄糖代谢方面,丁酸盐、乙酸盐、胆汁酸和吲哚刺激胰高血糖素样肽1的释放,从而间接影响胰岛素分泌和葡萄糖稳态。越来越多的证据还强调SCFA-G蛋白偶联受体(GPCR)轴在胰腺β细胞功能和胰岛素分泌调节中发挥关键作用。
铁可能在细菌衍生的SCFAs的调控中发挥作用。除了前面提到的对铁吸收的可能影响外,体内和体外模型显示出不同的铁-SCFA相互作用。例如,宿主缺铁对SCFA水平有很强的调节作用,尤其是丁酸盐。与喂食正常食物的大鼠相比,喂食缺铁饮食大鼠的丁酸盐和丙酸盐水平在统计学上显著降低,同时主要产生SCFA的物种减少。在体外结肠发酵研究中也观察到了类似的发现。在这些实验中,极低浓度的铁严重损害了SCFAs的产生并降低了丁酸盐生产菌的相对丰度。SCFAs中与铁相关的变化也可能与肠道微生物群组成的变化无关。SCFA生物合成途径包含几种铁依赖性酶,并且在体外模型中缺铁会下调参与丁酸盐生产的微生物群基因的表达。 在体外实验中,细菌LPS通过激活NF-κB来调节巨噬细胞和树突状细胞中的铁蛋白转录。微生物群产生的代谢物通过抑制十二指肠上皮细胞中HIF2α的表达和诱导铁储存蛋白铁蛋白的表达来限制铁的吸收,从而调节小鼠体内铁稳态。对微生物代谢物进行高通量筛选后,发现丁酸盐、丙酸盐和1,3-二氨基丙烷(DAP)在体外抑制三种人肠道细胞系中的HIF2α表达和活性,即使它们没有改变铁蛋白表达。但来自无特定病原体小鼠粪便的代谢物粗提物也诱导了这些细胞系中铁蛋白的表达,突出了其他未鉴定的肠道微生物代谢物在铁储存调节中的作用,与LPS无关。在这些代谢物中,只有DAP在两个已建立成熟的HIF2α诱导小鼠模型中抑制了体内肠道HIF2α活性。与DAP类似,在体外罗氏菌素(罗伊氏乳杆菌的一种微生物代谢物)在人肠上皮细胞系中具有HIF2α抑制特性。补充这些代谢物可减少血色素沉着症小鼠模型中的铁积累。抗生素治疗和微生物群定植无菌小鼠导致两种代谢物(DAP和罗氏菌素)水平的显著改变,表明微生物群是其水平升高的主要贡献者。 丙酸咪唑是一种组氨酸的微生物衍生代谢物,会损害肝脏、比目鱼肌和无菌小鼠的白色脂肪组织裂解物以及原发性肝细胞中的胰岛素信号。2021年在两个不同的队列中(一个由严重肥胖的个体组成,另一个由患有和无肥胖的健康个体组成),组氨酸是与血清铁蛋白浓度负相关性最强的血浆代谢物。在这些队列中,来自链球菌科、Pasteurellaceae和甲烷杆菌科的细菌在血清组氨酸水平下降和肝脏脂质积累的个体中富集。
6.2 减弱毒力的相互作用
有证据表明宿主-微生物组争夺铁的竞争模式发生了变化,强调了宿主-微生物组-铁协同相互作用在减弱微生物毒力和促进宿主存活方面的重要作用。例如,小鼠实验模型中的柠檬酸杆菌(Citrobacter rodentium)感染表明宿主铁代谢可能诱导协同防御机制,通过触发宿主生理变化来减弱病原体毒力并促进长期无症状感染。感染C. rodentium小鼠的肝脏转录组显示与未感染对照和死亡的感染小鼠相比,感染后存活的宿主中参与铁代谢的基因表达上调。膳食铁补充通过诱导胰岛素抵抗、减少肠道葡萄糖吸收和增加肠腔葡萄糖浓度来抑制C. rodentium毒力因子的表达。这种变化导致肠道中病原体的葡萄糖可用性增加,从而导致其毒力转录过程的抑制。当营养物质缺乏时,铁在降低微生物组的毒力方面至关重要。同样,对败血症的耐受性(在小鼠盲肠结扎和穿刺模型中)依赖于铁和葡萄糖代谢反应间的相互作用。铁蛋白可抵消肝葡萄糖-6-磷酸酶的铁血红素抑制,促进肝糖异生产生内源性葡萄糖,从而通过预防低血糖诱导对败血症的疾病耐受性。
血红素解毒作用构成了另一种宿主-微生物组协同抗毒机制,它通过减少细胞损伤来促进宿主生存和发育。宿主体内的血红素降解由HO1介导,它催化血红素分解为一氧化碳、胆绿素和铁。值得注意的是,HO1似乎是胰岛素抵抗的驱动因素。肝脏和脂肪组织HO1活性是小鼠和人类胰岛素抵抗最强的独立预测因子之一。小鼠肝脏和巨噬细胞特异性敲除HO1可提高胰岛素敏感性、减少炎症并减轻高脂饮食诱导的肝脏脂肪变性。
6.3 代谢相关性脂肪肝病
人体中血清铁蛋白水平与可能调节肝脏糖异生和脂质积累的特定微生物功能特征相关,并可能影响代谢相关性脂肪肝病(MAFLD)。严重肥胖患者的血清铁蛋白浓度与双歧杆菌、乳杆菌和韦荣球菌(Veillonella)丰度呈负相关,而与拟杆菌(Bacteroides)和普氏菌(Prevotella)呈正相关。据报道,人类队列中双歧杆菌、乳杆菌和韦荣球菌与MAFLD进展呈负相关,而普氏菌与MAFLD进展呈正相关。严重肥胖个体的血清铁蛋白相关微生物科也与肝脏中铁代谢相关基因的表达有关:血清铁蛋白水平与Pasteurellaceae、Leuconostocaceae和Micrococcaceae的丰度呈负相关,它们也与肝脏TFRC、铁调素、IRP1和FPN表达有关。
微生物组的功能也与血清铁蛋白水平有关。参与铁吸收的细菌细胞色素b561的功能与严重肥胖患者的血清铁蛋白水平呈负相关。在具有高血清铁蛋白水平的个体中,几种细菌的铁转运和螯合功能以及参与血红素和铁载体生物合成的细菌酶减少。宿主血清铁蛋白水平还与参与谷胱甘肽生物合成和谷胱甘肽前体的细菌功能有关,并与细菌甘氨酸转运蛋白和谷氨酸-半胱氨酸连接酶(谷胱甘肽生物合成途径中的第一种酶)呈负相关。血浆和肝脏谷胱甘肽水平是众所周知的MAFLD患者的关键特征。
6.4 铁和脂肪组织
铁还影响白色脂肪组织中的胰岛素抵抗。在C. rodentium感染期间,小鼠的膳食铁供应导致白色脂肪组织中铁水平和胰岛素抵抗增加。有趣的是,胰岛素抵抗的个体脂肪组织铁含量也增加。 2021年的一项研究揭示了脂肪细胞铁水平影响脂肪-肠道相互作用以调节肠道脂质吸收和介导肥胖保护作用。喂食高脂肪饮食的小鼠脂肪细胞特异性缺失Tfr1导致脂肪组织铁积累和饮食脂肪吸收减少,同时小肠炎症和氧化应激减少。肠道微生物群的健康组成变化,特别是变形菌门(Deltaproteobacteria、Epsilonprotebacteria和Gammaproteobacteria)的细菌减少,而Coriobacteria和Bacilli增加。一些研究还将微生物组与人体组织中的铁含量联系起来。有趣的是,在明显健康的肥胖和非肥胖个体中,微生物群中不同细菌铁载体的存在与中枢性肥胖和大脑铁含量的发展有关。
7 进化观点
铁氧化细菌(其中大部分属于变形菌门)对地球的地球化学演化具有重大影响。如前所述,铁可能塑造了生命的外观甚至人类的进化(图5)。在几种情况下,铁还在宿主-微生物的协同相互作用中发挥作用(Box 2)。 从人类进化的角度来看,胰岛素抵抗和铁代谢紊乱有着明显的悖论,两者在人群水平上都非常普遍,但与发病率和死亡率的增加有关。西方国家超过25%的人口受胰岛素抵抗的影响,而遗传性血色素沉着病是欧洲血统人群中最普遍的遗传疾病之一。欧洲人HFE中遗传性血色素沉着病C282Y的高频率突变已被认为是突变杂合携带者的生存优势。这种突变使其对通常感染巨噬细胞的微生物产生抵抗,如结核分枝杆菌(Mycobacterium tuberculosis)(引起肺结核)或伤寒沙门氏菌Salmonella enterica serovar Typhi(引起伤寒)。然而,遗传性血色素沉着症患者也极易感染Yersinia enterocolitica和Vibrio vulnificus。值得注意的是,考虑到在静脉切开术后遗传性血色素沉着病患者的肠道微生物组发生的重要变化以及相关的结肠铁水平降低,整体情况要复杂得多。 轻度全身炎症与从饮食中的铁摄取相互作用。例如,超重和肥胖女性的中枢性肥胖与血清铁调素水平升高有关,这导致补铁后吸收减少、全身铁利用率降低以及转铁蛋白饱和度降低。矛盾的是,肝脏铁积累也增加了。这些与肥胖相关的铁稳态紊乱被称为“代谢紊乱铁过载综合征”。实际上许多研究表明肥胖个体缺铁且转铁蛋白饱和度降低。然而其他研究报告了肥胖个体组织中铁(铁蛋白)标志物增多,红细胞生成(血红蛋白)的铁利用率增加。这些结果可能是由于铁向代谢组织的转移增加,这有利于全身炎症的增加。事实上,MRI研究显示严重肥胖患者肝脏、脂肪组织和骨骼肌中铁的积累增加。
组织铁积累会导致氧化应激和局部炎症增加,损害胰岛素依赖组织中的胰岛素信号传导并扰乱全身葡萄糖代谢。 导致循环先天免疫蛋白、细胞模式识别受体和炎症细胞因子的产生或功能改变的遗传变异与肥胖、胰岛素抵抗和T2DM相关。进化压力,例如人群水平的急性感染(大流行)和长期环境产物(LPS)或传染性病原体的低暴露,可能导致易感性以及目前的胰岛素抵抗和T2DM患病率增加。我们建议在竞争和合作层面将铁添加到这个等式中;在互惠相互作用中铁可以用来交换养分。
事实上,铁和葡萄糖代谢之间因果关系存在的证据很少。T2DM的一些特征,例如肝葡萄糖生成增加和代谢不灵活性,已在高脂饮食的血色素沉着症小鼠模型中进行了描述,这可能导致组织铁过多所致糖尿病的风险。肠道微生物组有助于探索铁和葡萄糖代谢之间的因果关系。
Box 2 通过铁的宿主-微生物协同相互作用。
有利于线粒体生物发生:细菌来源的肠杆菌素与小鼠宿主线粒体ATP合酶的α亚基结合,从而促进宿主铁的获取和生长。
促进伤口愈合:铁调素是铁代谢的关键因子,由2型常规树突状细胞产生,可促进肠道内稳态。常规树突状细胞特异性铁调素缺陷小鼠在肠道损伤后恢复速度较对照小鼠慢,这可能会影响肠道微生物群。
有助于解毒、循环和葡萄糖调控:铁蛋白诱导的肝脏糖异生对于多微生物败血症期间小鼠宿主存活至关重要。
图5 铁和葡萄糖获取如何推动人类进化的模型。
该图总结了关于饮食中铁和葡萄糖水平(即缺乏或过量)和宿主基因型(胰岛素抵抗与胰岛素敏感)之间相互作用的假设。进化结果集中在细菌感染的易感性和/或死亡率和存活率。
结论
体内稳态(葡萄糖和铁代谢)的调节需要考虑肠道微生物群在其与宿主的双向相互作用中的作用。免疫系统通过可溶性因子的产生和巨噬细胞的活性不断地感知和调节宿主-微生物群界面的相互作用。微生物群与宿主之间对铁的竞争和互利合作(降低细菌毒力)对宿主产生了深远的影响。使用个性化和特定的以疾病为导向的方法对这种相互作用进行深入分析,不仅对感染性疾病具有治疗意义,而且对代谢疾病中发生的慢性轻度炎症也具有治疗意义。探索体液中的微生物群及其代谢物以及其与胰岛素依赖组织生理学的关联将揭示新的疾病机制。最早的案例之一是肠道宏基因组学-肝脏转录组学-全身葡萄糖代谢轴,显示了微生物群组成和功能、肝脏转录组学、全身葡萄糖代谢和胰岛素敏感性之间的关联。我们旨在将这些知识转化为针对外周胰岛素抵抗的新方法,避免糖耐量受损的发展并改善T2DM患者的代谢控制。 迫切需要进一步研究铁代谢、微生物群和细胞生理学之间的交互作用。先天免疫系统的造血细胞和非造血细胞需要铁来维持生理技能,并战略性地位于宿主-微生物组界面。高达90%的结直肠癌风险被认为由饮食等环境因素引起,这与被称为致癌性结直肠癌相关的微生物群一致。然而,铁与癌症的关系不仅限于肠道,例如癌细胞似乎通过增强脂质运载蛋白2的表达与巨噬细胞竞争铁,从而在脑脊液中存活。
不感兴趣
看过了
取消

人点赞

人收藏

打赏
不感兴趣
看过了
取消
©2012-2023 北京华媒康讯信息技术股份有限公司 All Rights Reserved. 注册地址:北京 联系电话:010-82736610
广播电视节目制作经营许可证 —(京)字第 17437号 京海食药监械经营备20200522号
京ICP备12011723号 京ICP证150092号
 京公网安备 11010802020745号
工商备案公示信息
互联网药品信息服务资格证书((京)-非经营性-2020-0015)
京公网安备 11010802020745号
工商备案公示信息
互联网药品信息服务资格证书((京)-非经营性-2020-0015)






您已认证成功,可享专属会员优惠,买1年送3个月!
开通会员,资料、课程、直播、报告等海量内容免费看!

打赏金额
认可我就打赏我~
1元 5元 10元 20元 50元 其它
打赏作者
认可我就打赏我~
扫描二维码
立即打赏给Ta吧!
温馨提示:仅支持微信支付!
已收到您的咨询诉求 我们会尽快联系您




































 010-82736610
010-82736610
 股票代码: 872612
股票代码: 872612




