
仍然存在的一个问题是存在于小鼠肠道中的真菌铁载体的起源。本研究中使用了SPF实验室条件下饲养的小鼠。
编译:微科盟蔚蓝,编辑:微科盟居居、江舜尧。
微科盟原创微文,欢迎转发转载,转载须注明来源《微生态》公众号。
导读
肠道真菌菌群(mycobiota)与破坏肠道内稳态的疾病有关,如炎症性肠病。然而人们对感染性结肠炎期间肠道中细菌和真菌之间的功能关系知之甚少。本研究探索了真菌代谢产物在肠道病原体Salmonella enterica serovar Typhimurium感染(全球胃肠炎的主要病因)期间的作用。研究发现肠道内食物中存在的真菌菌群和真菌都可能是铁载体的来源,铁载体是一种从宿主中清除铁的小分子。在体外和小鼠模型中,利用真菌铁载体的能力使表达真菌铁载体受体FhuA或FhuE的沙门氏菌菌株具有竞争生长优势。本研究强调了真菌和沙门氏菌之间的跨界互养作用,并阐明了肠道真菌菌群的额外功能,揭示了这些未被充分研究的肠道生态系统成员在细菌感染期间的重要性。
论文ID
原名:Mycobiota and diet-derived fungal xenosiderophores promote Salmonella gastrointestinal colonization
译名:真菌菌群和饮食来源的真菌异种铁载体促进沙门氏菌胃肠道定植
期刊:Nature Microbiology
IF:30.964
发表时间:2022.11
通讯作者:Judith Behnsen
通讯作者单位:美国伊利诺伊大学芝加哥分校
DOI号:10.1038/s41564-022-01267-w
实验设计
结果
1.沙门氏菌利用真菌铁载体在体外生长
基因表达分析证实,编码铁载体受体IroN (沙门氏菌素受体)、FepA (肠杆菌素受体)、FhuA(高铁色素受体及相关铁载体受体)和FhuE的沙门氏菌基因在LB培养基中表达。在添加铁螯合剂(营养液(NB)+NBD))的培养基和DMEM/F12+10%胎牛血清(FBS)培养基中表达上调(图1a)。NBD用于在完全铁限制的环境中培养沙门氏菌,而DMEM/F12+10% FBS特异性使沙门氏菌利用铁载体生长,因为铁与血清蛋白结合。在缺乏铁的基本培养基中也发现了类似的真菌铁载体受体基因上调,因此该培养基以及DMEM/F12+10% FBS适用于本研究中的体外实验(图1a)。我们还使用链霉素预处理小鼠模型检测了沙门氏菌铁载体受体基因表达。在沙门氏菌感染48小时后收集的粪便样本中,与iroN相比,fhuE的转录水平相似,而fhuA的转录水平更高,而iroN之前被证明可以促进沙门氏菌的胃肠道定植(图1a)。因此,真菌铁载体受体基因的表达可能对沙门氏菌的致病机制非常重要。接下来研究者在体外测试了沙门氏菌通过真菌铁载体获得铁的能力。野生型(WT)沙门氏菌在DMEM/F12+10% FBS中生长良好,而缺乏受体IroN(ΔiroN)导致生长略有下降,因为突变体只能利用肠杆菌素(图1b)。添加宿主蛋白脂质运载蛋白2(Lcn2)(隔离肠杆菌素)后完全抑制ΔiroN突变体的生长(图1b)。基因fhuA和fhuE的缺失并没有导致额外的生长缺陷(图1b)。添加纳摩尔量的真菌铁载体高铁色素或coprogen(desferri形态)可完全恢复ΔiroN菌株的生长(图1c,d)。真菌铁载体不能促进缺乏FhuA和FhuE受体的菌株的生长,显示出受体特异性(图1c,d)。使用同时缺失fepA和iroN的沙门氏菌菌株也得到了类似的结果,它们不能吸收沙门氏菌产生的铁载体。添加高铁色素或coprogen均能完全恢复ΔiroNΔfepA菌株的生长。相比之下,coprogen和高铁色素并不能挽救缺少两种真菌铁载体受体的突变体(ΔiroNΔfepAΔfhuAΔfhuE)的生长。红酵母酸,一种真菌铁载体,之前被发现在Escherichia coli K12中与FhuE结合,不能恢复ΔiroNΔfepA突变体的生长。所有菌株在LB或加入通过FoxA受体吸收的细菌异种铁载体去铁胺时都可达到野生生长水平(图1e)。在指数生长阶段,真菌铁载体的存在显著促进了WT沙门氏菌的生长(图1f)。值得注意的是,高铁色素对WT沙门氏菌生长的促进作用强于coprogen。最后,WT沙门氏菌与ΔfhuAΔfhuE沙门氏菌在含有高铁色素的基本培养基中共培养时具有竞争优势。当向培养基中添加去铁胺时,并未发现竞争优势(图1g)。
图1 沙门氏菌能够利用纯化的真菌铁载体。
a,实时PCR分析在LB(n=3)、DMEM/F12+10% FBS(n=3)、NBD(n=3)、基本培养基(n=3)指数型增长的沙门氏菌培养物和感染WT沙门氏菌(n=4) 48h的常规饲养小鼠结肠腔内容物中的铁载体受体(iroN、fepA、fhuA和fhuE)编码基因。b-d,接种后8小时DMEM/F12+10% FBS和DMEM/F12+10% FBS+100 ng ml−1脂质运载蛋白2(WT n=5; iroN n=5(DMEM) n=6 (DMEM+Lcn2);iroNfhuA n=3(DMEM) n=4(DMEM+Lcn2);iroNfhuE n=3)(b),DMEM/F12+10% FBS+100 ng ml−1脂质运载蛋白2+100 nM coprogen (n=3)(c),DMEM/F12+10% FBS+100ng ml−1脂质运载蛋白2+100nM高铁色素(n=3)(d)中沙门氏菌菌株WT、iroN、iroNfhuA和iroNfhuE的生长。e,接种后8小时DMEM/F12+10% FBS(n=10);DMEM/F12+10% FBS+100 nM高铁色素(n=3);DMEM/F12+10% FBS+100 nM coprogen(n=3);DMEM/F12+10% FBS+100 ng ml−1去铁胺(n=4);DMEM/F12+10% FBS+100 ng ml−1红酵母酸(n=3)中沙门氏菌菌株WT、iroNfepA和iroNfepAfhuAfhuE的生长;f, WT沙门氏菌在基本培养基、基本培养基+100 nM高铁色素、基本培养基+100 nM coprogen(n=3)中的生长。g,在基本培养基(n=3)、基本培养基+100 nM高铁色素(n=3)、基本培养基+100 ng ml−1去铁胺(n=3)中WT沙门氏菌/fhuAfhuE的竞争指数(输出比(CFU ml−1iroNfepA)/(CFU ml−1iroNfepAfhuAfhuE)除以输入比)。a-g中数据用平均值±标准差(s.d.)表示。统计学:普通单因素方差分析(ANOVA)与Tukey多重比较检验(a, c和d);双因素方差分析与Tukey多重比较检验(b和e-g)。P值在图中显示。
2.肠道真菌产生的铁载体促进沙门氏菌的生长
随后我们调查了肠道真菌微生物组中是否存在产生铁载体的真菌。我们对从C57BL/6小鼠粪便样本中提取的DNA进行了内部转录间隔区(ITS)测序。测序结果确实揭示了一些真菌属具有已知的产生铁载体的物种,例如曲霉属Aspergillus和青霉菌属Penicillium(图2a)。我们选择了四种真菌并在体外检测其促进沙门氏菌生长的能力。黑曲霉(Aspergillus niger)是曲霉属的代表成员,Malassezia furfur是人类皮肤和胃肠道的共生菌。我们还检测了一种从本研究小鼠中分离出来的丝状真菌,它可能是Scopulariopsis属的一个成员(称为小鼠分离株),也包含已知的铁载体生产菌。我们与能够(ΔiroNΔfepA)和不能(ΔiroNΔfepAΔfhuAΔfhuE)利用每种真菌存在或不存在的真菌铁载体的沙门氏菌菌株进行了竞争性体外共培养。在没有真菌的情况下,两种沙门氏菌菌株在铁限制的基本培养基中生长相似(竞争指数1)(图2b)。所有被检测的真菌物种都能在这种培养基中生长。在产生铁载体的真菌存在的情况下(即A. niger、M. furfur和小鼠分离株),能够利用真菌铁载体的沙门氏菌菌株(ΔiroNΔfepA)的生长速度平均提高了15倍(图2b)。另一方面,酵母Saccharomyces cerevisiae的存在不能产生铁载体,没有提供任何竞争优势(图2b)。从培养A. niger和小鼠分离株72小时的基本培养基中获得的无菌过滤上清也具有竞争优势,因此表明真菌铁载体被释放到培养基中(图2c)。
许多真菌也利用铁载体在细胞内储存铁。因此,我们测试了细胞内真菌铁载体(结构上与分泌铁载体不同)是否也促进了沙门氏菌的生长。丝状真菌A. niger和小鼠分离株在富培养基(酵母浸出粉胨葡萄糖,YPD)中生长,在基本培养基中清洗和均质。在经无菌过滤的真菌匀浆中生长8小时后,沙门氏菌ΔiroNΔfepA的生长速度较不能吸收真菌铁载体的菌株提升10倍(图2d)。由于沙门氏菌可以杀死真菌,这一发现表明沙门氏菌可能具有获得真菌细胞内容物的能力。因此,沙门氏菌可以利用纯化的真菌铁载体、交叉喂养期间真菌提供的铁载体和细胞内真菌铁载体作为铁源。
图2 肠道真菌是真菌铁载体的来源。
在未感染SPF小鼠的粪便样本中利用ITS测序鉴定的真菌属的相对丰度(n=6)。b-d,在基本培养基添加活真菌(b)、真菌上清液(n=3)(c)及真菌匀浆(n=3)(d) 8h后沙门氏菌菌株iroNfepA对iroNfepAfhuAfhuE的竞争指数(n=4)。b-d中数据用平均值±s.d.表示。统计:普通单因素方差分析与Dunnett多重比较检验。P值在图中显示。
3.饮食变化深刻地改变了肠道真菌菌群的组成
在铁限制条件下铁载体的吸收很重要。然而,实验室小鼠通常被喂食的饮食中铁含量为200-350 ppm,远超过了它们的饮食需求35 ppm,而且远高于人类标准饮食中的铁含量。为了测试体内真菌铁载体的作用,我们使用了含有50 ppm铁的精制饲料。由于以谷物为基础的常规饲料和精制饲料配方差异显著,我们评估了它们对细菌和真菌菌群的影响。我们给小鼠喂食谷物为基础的常规饲料(饲料LM485,铁含量为240 ppm) 1周,然后再给小鼠喂食含50 ppm铁的精制饲料1周(图3a)。ITS和16S测序显示细菌和真菌种群以饮食为基础聚集。更换饲料后,细菌α多样性降低(图3b),大多数相关细菌属的相对丰度发生改变(图3c)。更换饲料后,真菌微生物组发生了最显著的变化,α多样性显著降低(图3b),真菌属的相对丰度发生了深刻变化(图3d)。喂食常规饲料LM485时大多数小鼠定植曲霉属、Cladosporium和青霉菌属真菌,如图2a所示。然而,在喂食精制饲料1周后,即使是同屋饲养的小鼠,真菌群落组成也具有显著的个体间变异。考虑到精制饲料对肠道菌群中存在的菌属的显著影响,我们测试了换为另一种以谷物为基础的常规饲料(饲料2914)是否也会影响肠道真菌微生物群(图3e)。细菌微生物组的α多样性(图3f)没有显著变化,主成分分析显示重叠聚类。最具代表性的细菌属的相对丰度保持相似性(图3g)。与更换为精制饲料相比,真菌微生物组组成的变化也不太明显。然而,真菌微生物组仍然以饲料为基础聚类,α多样性降低(图3f)。在所有被测试的小鼠中,最具代表性的属是曲霉菌属,而在单个小鼠中,没有单一的属或物种的比例过高(图3h)。因此,饮食可能直接调节真菌微生物组的组成,或其本身是小鼠肠道中真菌序列的来源。事实上,之前的研究表明,不同的饲料可能含有真菌DNA。因此,我们测试了不同小鼠饮食配方中真菌DNA的数量。我们分析了两种以谷物为基础的常规饲料(LM485和5066(类似于2914))以及两种不同铁浓度(50 ppm和200 ppm)的精制饲料。在小鼠常规饲料中检测到大量的真菌DNA,但在精制饲料中检测到的水平明显较低(图3i)。因此,真菌DNA可以通过以谷物为基础的常规饲料进入肠道环境。
图3 饮食变化深刻地改变了肠道真菌微生物组的组成。
a,测序分析实验装置示意图。FS,粪便样本;b, α多样性分析,16S(左)和ITS(右)的Chao指数(n=5);c、d,改变饮食前后小鼠粪便样本中经测序鉴定的细菌(c)和真菌(d)属的平均相对丰度(n=5);e,测序分析的实验设置示意图。f, α多样性分析,16S(左)和ITS(右)的Chao指数(n=5)。g、h,改变饮食前后小鼠粪便样本中经测序鉴定的细菌(g)和真菌(h)属的相对丰度(n=5);i,不同小鼠饮食中真菌DNA的实时PCR分析。(i)(常规饲料LM485 n=5, 常规饲料5066 n=6,精制饲料50 ppm n=5,精制饲料200 ppm n=4)。统计学:非配对t检验(b和f);普通单因素方差分析与Tukey多重比较检验(i)。P值在图中显示。
4.饮食中真菌铁载体促进沙门氏菌生长
当我们在小鼠常规饲料中检测到相对热不稳定的真菌DNA(图3i)时,我们推测热稳定的化合物可能也存在,如真菌铁载体。为了测试铁载体是否存在于小鼠常规饲料中,我们使用了生物指示剂Microbacterium flavescens JG-9菌株,这是一种异羟肟酸型铁载体营养缺陷菌。异羟肟酸型铁载体是真菌产生的铁载体的主要种类,高铁色素和coprogen都属于这一类。使用生物指示剂是必要的,因为通过质谱法对铁载体的鉴定和定量仅能在液体和最小复杂的样品中成功,如尿液,而在复杂的悬浮液中(即饮食或粪便)不能实现这一点。M. flavescens在添加异羟肟酸型铁载体去铁胺的LB培养基上生长良好(阳性对照),在未添加它的情况下无法生长(图4a)。用常规饲料5066制成的混悬液也促进了M. flavescens的生长。相比之下,精制饲料(含50 ppm和200 ppm铁)和常规饲料LM485制成的悬浮液不支持M. flavescens的生长(图4a)。因此,一些以谷物为基础的小鼠常规饲料可能含有异羟肟酸型铁载体。这一点在沙门氏菌菌株之间的竞争生长试验中被证实。在以谷物为基础的常规饲料中,沙门氏菌ΔiroNΔfepA的竞争力是ΔiroNΔfepAΔfhuAΔfhuE的5倍(LM485)到10倍(饲料5066)。常规饲料LM485为ΔiroNΔfepA沙门氏菌提供了竞争优势,表明除M. flavescens可利用的铁载体外,这种常规饲料中还存在其他铁载体。相比之下,无论总铁含量如何,精制饲料并未带来竞争优势(图4b)。在精制饲料悬液中添加高铁色素后,ΔiroNΔfepA菌株的竞争优势恢复到类似于饲料5066的水平(图4c)。
在更接近肠道环境的厌氧环境中,以谷物为基础的饲料也为ΔiroNΔfepA提供了竞争优势(图4d)。与需氧振荡培养相比,其竞争优势较低(图4b),可能是由于有限的营养物质混合物,因为需氧非振荡液培养与振荡培养相比也显示出较低的竞争指数(图4d)。最后,我们想评估人类食物中是否含有真菌铁载体并促进沙门氏菌生长。为此,我们在过滤均质悬浮液中对沙门氏菌菌株进行共培养。以谷物为基础的人类食物,如面粉和白面包,赋予了沙门氏菌菌株ΔiroNΔfepA竞争优势(图4e)。含有青霉菌属活真菌的蓝奶酪是已知的铁载体生产者,使ΔiroNΔfepA拥有ΔiroNΔfepAΔfhuAΔfhuE 60倍的竞争优势。同样,由Fusarium组成的肉类替代品Quorn也产生铁载体,使ΔiroNΔfepA的生长比ΔiroNΔfepAΔfhuAΔfhuE快17倍。不太可能含有真菌的食物(香蕉和鸡肉),以及不产生铁载体的酵母发酵产物(啤酒和葡萄酒)并没有为ΔiroNΔfepA沙门氏菌提供竞争优势。这些结果与我们的假设一致,即真菌铁载体可以通过饮食摄入进入小鼠和人体的肠道环境。
图4 小鼠和人类的饮食中可能含有促进沙门氏菌生长的真菌铁载体。
a,在添加不同饲料悬液、35.6 μM去铁胺的LB或单用LB中,接种96h后M. flavescens的生长情况。b,在有氧振荡液体培养中添加饲料悬液后,沙门氏菌菌株iroNfepA对iroNfepAfhuAfhuE的竞争指数。c,在添加饲料悬液和降低高铁素浓度的基本培养基中,接种8h后沙门氏菌菌株iroNfepA对iroNfepAfhuAfhuE的竞争指数。d,在添加饲料悬液的基本培养基中需氧非振荡液体培养(左)和厌氧非振荡液体培养(右),接种8h后沙门氏菌菌株iroNfepA对iroNfepAfhuAfhuE的竞争指数。e,在添加人类食物悬液的基本培养基中,接种8h后沙门氏菌菌株iroNfepA对iroNfepAfhuAfhuE的竞争指数。在a-e中,数据表示为平均值±s.d.。统计学:采用单因素方差分析和Dunnett (a, c和e)或Tukey (b和d)多重比较检验。
5.真菌铁载体促进沙门氏菌肠道生长
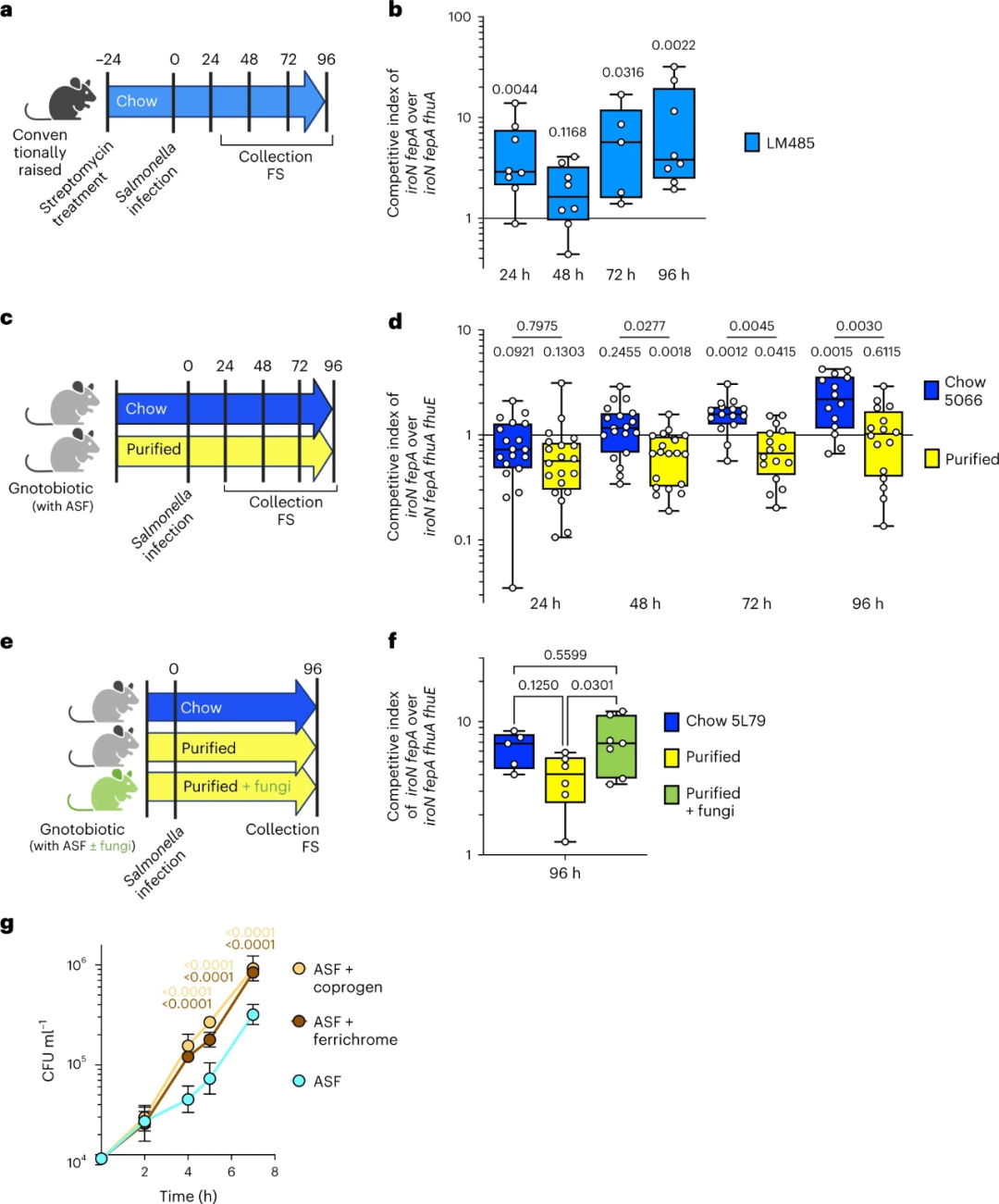
本研究最终想要确定真菌铁载体是否在体内沙门氏菌感染中发挥作用。研究中使用的是具有完整真菌和细菌微生物群的常规饲养的特异性无病原体(SPF)小鼠。精制饲料对细菌和真菌微生物群组成有较深影响(图3c,d),我们使用含有真菌铁载体的常规饲料LM485喂养小鼠(图4b,c)。用链霉素处理小鼠,使沙门氏菌诱导结肠炎,24h后以1:1比例的沙门氏菌菌株ΔiroNΔfepA和ΔiroNΔfepAΔfhuA感染小鼠。每天收集粪便样本,并在感染后96小时处死小鼠(图5a)。沙门氏菌ΔiroNΔfepA引起与WT沙门氏菌水平相似的炎症,并且能够在粪便样本的所有时间点上战胜ΔiroNΔfepAΔfhuA(图5b)。在Peyer斑(P=0.0657)和肠系膜淋巴结(P=0.0033)中也发现了竞争优势。为区分饮食和肠道共生真菌提供的真菌铁载体,我们利用特定共生菌群组合(ASF)定植的定菌小鼠进行了两个独立的实验。ASF是由8种细菌组成的菌群,因此,ASF定植的小鼠将缺乏真菌,但仍有细菌作为铁的竞争者。我们通过给ASF小鼠喂食精制饲料、常规饲料5066或饲料5L79来分析饲料的作用。因为与含有50 ppm和200 ppm铁的饲料的体外共培养结果相似(图4b,d),我们选择使用铁含量为200 ppm,这个铁含量更接近于常规饲料5066和饲料5L79。小鼠口服1:1比例的ΔiroNΔfepA和ΔiroNΔfepAΔfhuAΔfhuE沙门氏菌菌株,并每天收集粪便样本(图5c)。与我们的体外实验结果一致,沙门氏菌菌株ΔiroNΔfepA在喂食以谷物为基础常规饲料的小鼠的肠道中具有竞争优势,但在喂食精制饲料的小鼠中未体现出竞争优势(图5d)。在喂食以谷物为基础的常规饲料的小鼠中,竞争优势随着时间的推移而增加(图5d),并在感染后72小时和96小时达到显著性(图5d)。在饲喂精制饲料的小鼠中,随着时间的推移,一些小鼠的竞争优势略有增加(图5d),但总体而言,在任何时间点都没有观察到菌株ΔiroNΔfepA的竞争优势(图5d)。
为测试共生真菌提供的铁载体的作用,我们通过喂养含铁载体生产者A. niger和M. furfur的200 ppm精制饲料将其定植于ASF小鼠中(图5e)。作为对照组,另外两组小鼠分别饲喂以谷物为基础的饲料5L79或不添加真菌的精制饲料。将1:1比例的ΔiroNΔfepA和ΔiroNΔfepAΔfhuAΔfhuE沙门氏菌感染小鼠,感染96h后采集粪便标本。与之前的结果一致,与饲喂精制饲料的小鼠相比,喂养以谷物为基础的常规饲料的小鼠中ΔiroNΔfepA沙门氏菌具有更高的竞争优势。与饲喂无真菌定植的精制饲料的小鼠相比,添加活真菌显著增加了ΔiroNΔfepA沙门氏菌的竞争优势(图5f)。
最后,我们想测试在与其他细菌竞争时真菌铁载体是否会促进WT沙门氏菌的生长。在此情况下,沙门氏菌能够利用自身的铁载体沙门菌素和肠杆菌素。我们在有或无真菌铁载体的基本培养基中进行沙门氏菌和ASF细菌的共培养。如图5g所示,即使在培养基中存在其他细菌的情况下,沙门氏菌在真菌铁载体存在的情况下生长得更好。这些结果证实了真菌铁载体对沙门氏菌摄取铁的重要性。总之,我们的数据表明沙门氏菌已经进化出一种在铁限制条件下(如肠道炎症)利用真菌铁载体作为额外铁来源的机制。
图5 在体内真菌铁载体赋予沙门氏菌竞争优势。
a,SPF小鼠实验装置示意图。FS,粪便样本。b, 24h(n=8)、48h (n=8)、72h (n=5)和96h (n=8)时SPF小鼠粪便样本中沙门氏菌菌株iroNfepA对iroNfepAfhuA的竞争指数。c, ASF小鼠实验设置示意图。d, 24h(常规饲料n=19,精制饲料n=20)、48h(常规饲料n=19,精制饲料n=20)、72h(常规饲料n=15,精制饲料n=16)和96h(常规饲料n=14,精制饲料n=16)时ASF小鼠粪便样品中沙门氏菌菌株iroNfepA对iroNfepAfhuAfhuE的竞争指数。e, 真菌定植ASF小鼠的实验装置示意图。f, 96h(常规饲料n=5, 精制饲料n=6, 精制饲料+真菌n=7)时ASF小鼠粪便样本沙门氏菌菌株iroNfepA对iroNfepAfhuAfhuE的竞争指数。g, 在添加ASF的基本培养基、基本培养基+100 nM高铁色素、基本培养基+100 nM coprogen中WT沙门氏菌的生长(n=3)。
讨论
铁和锌等微量元素对几乎所有形式的生命都至关重要。尤其是铁,在哺乳动物肠道中竞争激烈。在炎症条件下,肠道中铁的可用性减少,使致病菌饥饿致死,这一机制被称为营养免疫。细菌和真菌产生铁载体来获取这种金属元素,并在缺铁的环境中生存。最近,Pierce等人展示了大肠杆菌如何利用从干酪皮中分离出来的真菌产生的铁载体。Zhu及其团队也在体内证明了铁载体的交叉喂养,证明了多形拟杆菌(Bacteroides thetaiotaomicron)可以利用沙门氏菌产生的铁载体进而更好地在肠道生态系统中定植。
本研究的数据表明,真菌和沙门氏菌之间存在跨界铁载体互养,沙门氏菌可以通过受体FhuA和FhuE吸收真菌的铁载体。真菌铁载体如高铁色素和coprogen不能与宿主脂质运载蛋白-2结合,因此是病原体的额外铁源。本研究使用ΔiroNΔfepA背景的沙门氏菌菌株进行体内感染。这些菌株不能吸收沙门氏菌铁载体肠杆菌素和沙门氏菌素。因此,在炎症过程中产生的脂质运载蛋白-2的存在或缺失不会影响沙门氏菌利用肠杆菌素的能力。然而,当小鼠炎症水平升高时,能够利用真菌铁载体的菌株的竞争优势增加(图5d),这可能是由于在沙门氏菌引起的炎症期间肠道铁浓度总体较低所致。在体内WT沙门氏菌对真菌铁载体的吸收可能是多余的,对病原体的定植和传播影响有限。然而本研究表明沙门氏菌已经进化并保持了在体外限铁条件下利用铁载体作为潜在铁源的能力(图1b-g),且在需要时也可以在体内利用它们(图5b,d,f)。因此,利用真菌铁载体的能力可能是沙门氏菌在铁含量有限的环境中可以利用的工具。最后,我们强调了不同的饮食可以影响真菌微生物群的组成(图3d,h),并可能是真菌DNA的来源(图3i)。因此,科学家指出研究中使用的饮食类型是很重要的,因为不同的饮食可能会导致对结果的误解。
关于饮食和粪便中铁载体定量这一问题或未尝试过或未成功,目前的文献没有提供这类数据。如果能克服这一技术限制,研究人员就可直接研究铁载体在肠道中的作用。本研究探讨了真菌通过产生真菌铁载体来喂养沙门氏菌的能力。研究还表明,某些真菌物种(如酵母菌S. cerevisiae和Aspergillus nidulans)可以吸收和利用细菌铁载体,如肠杆菌素。综上所述,这些结果强调了铁载体在肠道生态系统中作为细菌和真菌菌群之间潜在的关键相互作用的重要性。
仍然存在的一个问题是存在于小鼠肠道中的真菌铁载体的起源。本研究中使用了SPF实验室条件下饲养的小鼠。在不太标准化的条件下,原位产生的真菌铁载体的重要性可能更大。在室外环境中饲养的小鼠肠道真菌含量丰富。值得注意的是,从这些小鼠的粪便中培养出了更多的曲霉菌属物种。曲霉菌的增加可能是由于食用了含有真菌孢子的食物。饮食也会影响人体肠道真菌菌群的组成。在一项小规模研究中,食用面包与粪便中鉴定的S. cerevisiae有关。本研究表明,可能含有活真菌或真菌污染物的人类食物促进了能够利用真菌铁载体的沙门氏菌的生长(图4e)。我们发现含有活真菌的奶酪(即蓝奶酪)在获得铁载体的基础上对沙门氏菌生长的影响最大。值得注意的是,奶酪是不同沙门氏菌爆发的载体。本研究结果还表明,不仅活真菌,真菌铁载体也可以通过饮食获得。这些结果增加了更多的病理条件,其中真菌的作用已体现出来。肠道环境中细菌和真菌之间新的相互作用正在不断地被挖掘。了解其性质和机制对于制定抗感染的替代策略至关重要。旨在减少活真菌和真菌铁载体摄入量的饮食干预可能是降低人类患者沙门氏菌定植和感染水平的潜在策略之一。
不感兴趣
看过了
取消

人点赞

人收藏

打赏
不感兴趣
看过了
取消
©2012-2023 北京华媒康讯信息技术股份有限公司 All Rights Reserved. 注册地址:北京 联系电话:010-82736610
广播电视节目制作经营许可证 —(京)字第 17437号 京海食药监械经营备20200522号
京ICP备12011723号 京ICP证150092号
 京公网安备 11010802020745号
工商备案公示信息
互联网药品信息服务资格证书((京)-非经营性-2020-0015)
京公网安备 11010802020745号
工商备案公示信息
互联网药品信息服务资格证书((京)-非经营性-2020-0015)






您已认证成功,可享专属会员优惠,买1年送3个月!
开通会员,资料、课程、直播、报告等海量内容免费看!

打赏金额
认可我就打赏我~
1元 5元 10元 20元 50元 其它
打赏作者
认可我就打赏我~
扫描二维码
立即打赏给Ta吧!
温馨提示:仅支持微信支付!
已收到您的咨询诉求 我们会尽快联系您





































 010-82736610
010-82736610
 股票代码: 872612
股票代码: 872612




