

【关注】深度||溶酶体靶向降解技术新进展



来源:医药学术
前 言
靶向蛋白降解(targeted protein degradation, TPD)是近几十年里一项新兴的蛋白质降解技术,引起了化学生物学和药物发现领域极大的关注。根据蛋白降解途径不同,可分为溶酶体系统和蛋白酶体途径。由于溶酶体降解蛋白途径的降解对象涵盖了蛋白质、蛋白质多聚体、细胞器等,并且能够降解胞内和胞外的蛋白质。因此,基于溶酶体的生物物质选择性降解策略具有巨大潜力,基于溶酶体的降解剂开发将是通过蛋白质降解实现疾病治疗的重要方向。本文综述了近年来出现的新型的基于溶酶体途径的靶向蛋白质降解技术,主要包括溶酶体靶向嵌合体(lysosome-targeting chimera, LYTAC)技术、自噬靶向嵌合体(autophagytargeting chimera, AUTAC)技术、自噬体偶联化合物(autophagosome-tethering compound, ATTEC)技术以及分子伴侣介导的自噬(chaperone-mediated autophagy, CMA)嵌合体技术,分析了其优势和目前面临的挑战,对未来的药物发现提出了展望。
作 者 | GQW
01背景介绍
溶酶体系统(the lysosomal system)和蛋白酶体途径(the proteasome pathway)是细胞内两个最重要的蛋白降解途径[1],两种途径既相互独立又彼此联系,保证了细胞内物质的特异性降解。需要特别指出的是,溶酶体途径在调节细胞内外蛋白质稳态中发挥着重要的作用,这一点在后续会做进一步说明。
近年来,人们研究最多的是基于泛素化-蛋白酶体降解系统(the ubiquitin-proteasome system,UPS)的蛋白水解靶向嵌合体技术(proteolysis targeting chimeras,PROTAC)和分子胶技术(molecular glue)。其中,蛋白水解靶向嵌合体技术利用双功能分子通过一个连接链(linker)将目标蛋白的配体和E3泛素连接酶的招募配体相连接,进而招募E3泛素连接酶对目标蛋白进行泛素化,之后蛋白酶体会识别泛素标记的第五蛋白并将其进行降解。目前,ARV-110、ARV-471和CC-94676三种PROTAC分子已进入临床试验。而分子胶是一种通过形成三元复合物促进两种蛋白质的二聚化或共定位,从而产生多种生物学和药理学功能;诱导E3泛素连接酶底物受体与靶蛋白之间新型相互作用,从而导致靶蛋白降解的小分子。据估计,分子胶可以促进大约600种人类E连接酶与20000多种潜在的人类靶蛋白之间的相互作用,为探索新靶点和潜在的小分子药物提供了丰富的资源。但是分子胶的研究仍处于起步阶段,将分子胶开发成治疗剂还有很长的路要走[2]。
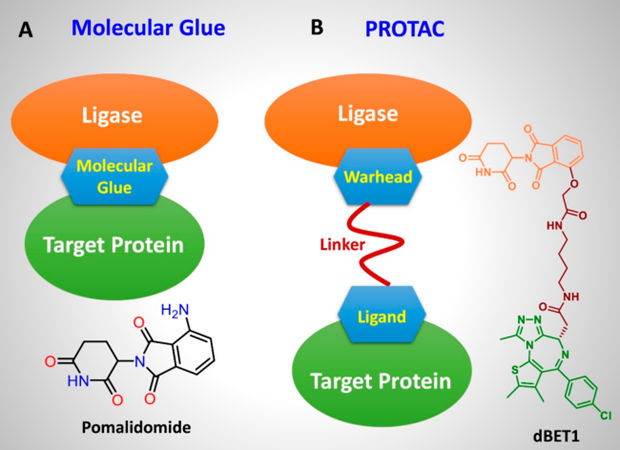
【图1】分子胶和PROTAC的作用方式及结构特点[2]
但是,上述两种基于蛋白酶体途径降解的技术只能降解某些细胞内蛋白质,并且由于需要依赖特定的E3连接酶和蛋白酶体,在某些细胞或具有蛋白酶体抗性的蛋白降解过程中,PROTAC和分子胶并不适用[3]。如前所述,由于溶酶体降解蛋白途径降解对象涵盖了蛋白质、蛋白质多聚体、细胞器等,并且能够降解胞内和胞外的蛋白质。因此,基于溶酶体的生物物质选择性降解策略具有巨大潜力,基于溶酶体的降解剂开发将是通过蛋白质降解实现疾病治疗的重要方向。
近年来,出现了许多新型的基于溶酶体途径的靶向蛋白质降解技术,主要包括溶酶体靶向嵌合体(lysosome-targeting chimera, LYTAC)技术、自噬靶向嵌合体(autophagytargeting chimera, AUTAC)技术、自噬体偶联化合物(autophagosome-tethering compound, ATTEC)技术以及分子伴侣介导的自噬(chaperone-mediated autophagy, CMA)嵌合体技术。
02 通过溶酶体途径降解蛋白质和细胞器
在人体内环境中,泛素-蛋白酶体系统主要降解可溶性、短寿命和错误折叠的单体蛋白,溶酶体系统主要降解长寿命蛋白、蛋白聚集体和受损的细胞器。通过溶酶体途径的蛋白质降解极大地扩展了已知底物范围,为靶向传统的非靶标蛋白质(如转录因子、细菌、病毒等)提供了一种新策略。
溶酶体膜上有介导物质运输出入的相关蛋白分布,蛋白质向溶酶体的转运需要溶酶体靶向受体(lysosome-targeting receptors, LTRs)参与。溶酶体的形成是一个复杂的过程,不同类型的细胞(甚至同一类型的细胞)可能采用不同的途径[4]。溶酶体水解酶在内质网中合成,主要通过以下两种方式进入溶酶体:
(1)首先是目前研究相对成熟的方法,即大多数溶酶体水解酶能够被甘露糖-6-磷酸受体(mannose-6-phospate receptor, M6PR)修饰,被高尔基体识别后转运至溶酶体[5];
(2)在另一种转运机制中,溶酶体整合膜蛋白2(lysosomal integral membrane protein 2, LIMP-2)介导葡萄糖脑苷脂酶(glucocerebrosidase, GCase)向溶酶体的转运[6](图2)。

【图2】溶酶体靶向降解生物物质的途径[6]
在组织中广泛表达的M6PR可以有效地将底物转运至溶酶体,因此可以筛选与M6PR、LIMP-2以及其他LTRs具有高亲和力的分子,并将其与靶蛋白链接,实现靶蛋白降解,从而开发靶向溶酶体降解技术。
溶酶体主要通过内吞作用和自噬两种方式选择性清除细胞内的蛋白聚集体、病原体和多余(或者受损)细胞器[8]。内吞作用始于质膜,止于溶酶体,是细胞吸收营养物质、与外部进行信息交换和更新质膜的重要途径;自噬是一种高度保守的细胞降解过程,可分为巨自噬(macroautophagy)、微自噬(microautophagy)和分子伴侣介导的自噬(chaperone-mediated autophagy, CMA)。
巨自噬是细胞内主要的自噬通路,在巨自噬过程中,自噬受体能够(如p62/SQSTM1)识别聚集的蛋白质并将其选择性封闭在自噬体内,接着在溶酶体水解酶作用下,自噬体与溶酶体融合,形成自噬溶酶体进而完成降解[9]。
微自噬是溶酶体或液泡内膜直接内陷将细胞内物质包裹并降解的过程。
分子伴侣介导的自噬(CMA)是指在含有可被溶酶体膜受体 LAMP-2A(溶酶体相关膜蛋白-2A)识别的胞浆蛋白的复合体内,胞浆蛋白在溶酶体膜上进行直接转运,使其去折叠并完成降解。这是一种独特的、具有高度选择性的自噬过程,只能降解某些特定的蛋白质,不能降解细胞器。
自噬过程在所有真核细胞中都存在,并且该过程在癌症、神经退行性疾病和代谢性疾病等许多疾病中异常进行[10]。因此,通过小分子诱导溶酶体发生自噬,从而降解某些靶蛋白,可能在药物发现领域有潜在的应用。
03 基于溶酶体途径的靶向蛋白质降解技术
1. 溶酶体靶向嵌合体(lysosome-targeting chimera, LYTAC)技术
溶酶体靶向嵌合体(LYTAC)技术是一种利用内吞-溶酶体途径降解目标蛋白的方法,主要作用于膜蛋白和细胞外对具有蛋白酶体抗性(此时PROTAC技术不再适用)的蛋白质(如EGFR)。
2020年7月,Banik等人[11]开发了第一个LYTAC:能够在细胞外介导TPD的甘露糖-6-膦酸酯(mannose-6-phosphonate, M6Pn)-抗体偶联物。M6Pn-LYTACs
连接了阳离子非依赖型甘露糖-6-磷酸受体(cation-independent mannose-6-phosphonate receptor, CI-M6PR, 一种溶酶体靶向受体LTR)和蛋白质靶标。形成的三元复合物经网格蛋白介导内吞进入细胞内,并经囊泡运送到早期内体(膜包裹的囊泡结构)中;随着内体酸化,CI-M6PR解离并回到细胞膜或高尔基体,LYTAC与靶蛋白的复合物被继续转运至溶酶体中降解。M6Pn被证明可以有效降解一系列靶标蛋白(如APOE4, EGFR, PD-L1, CD71等),但是由于CI-M6PR在细胞中普遍表达,关于M6Pn-LYTAC的适用性、安全性仍有待商榷。

【图3】LYTAC技术降解靶标蛋白质的过程[11]
2021年,三项相关研究揭示了靶向肝脏组织特异性去唾液酸糖蛋白受体(liver-specific asialoglycoprotein receptor, ASGPR)的N-乙酰半乳糖胺三联体(triantennary N-acetylgalactosamine, tri-GalNAc)靶向诱导相关蛋白的溶酶体降解。Ahn等人[12]和Zhou等人[13]均报道了基于FDA批准的EGFR抗体西妥昔单抗(cetuximab, Ctx)开发的GalNAc-LYTAC,顺利诱导了肝癌细胞系(hepatocellular carcinoma cell lines, HCC)的细胞表面EGFR受体的降解。

【图4】Ctx-GalNAc促进HCC细胞系表面EGFR的降解过程[12]
Ahn等人合成的HER2抗体帕妥珠单抗(petuzumab, Ptz, 用于治疗HER2阳性乳腺癌)-GalNAc复合物、多特异性整合素结合肽(polyspecific integrin binding peptide, PIP)-GalNAc复合物,分别顺利诱导了HEPG2上HER2以及细胞表面整合素的降解。其中,PIP-GalNAc是第一个非抗体LYTAC复合物。
Zhou等人通过将tri-GalNAc与生物素偶联,发现该复合物能够促进肝细胞通过ASGPR摄取中性亲和素蛋白(NeutrAvidin)并将其转运至溶酶体发生降解,证实了小分子LYTAC的可行性,为后续Caianiello等人的研究提供了参考。

【图5】Zhou等人对LYTAC的研究结果[13]
数月后,Caianiello等人[14]报道了MoDE-As,即靶向ASGPR诱导细胞外蛋白质降解的小分子降解剂,这是第一个由化学合成得到的双功能小分子与GalNAc三联体的复合物。他们发现的类PROTAC、异双功能化合物D-MoDE-A和M-MoDE-A,能够分别高效地诱导HepG2细胞中抗二硝基苯酚抗体(anti-dinitrophenol, α-DNP)和巨噬细胞游走抑制因子(macrophage migratory inhibitory factor, MIF)的降解,首次证明了非蛋白质的小分子化合物在体外和体内均能够有效介导细胞外蛋白质的降解,显示出LYTAC技术中小分子介导蛋白质降解的巨大潜力。同时,由于小分子更容易进行结构优化,因此可能更容易得到具有良好药代动力学和口服生物利用度的LYTAC复合物。
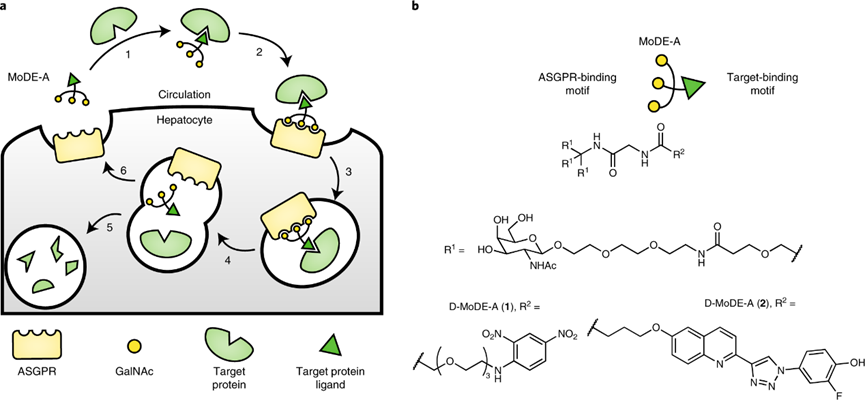
【图6】D-MoDE-A和M-MoDE-A的分子结构及其作用机制[14]
如上,目前已报道的与LYTAC相关的LTR仅有两种:阳离子非依赖型甘露糖-6-磷酸受体(CI-M6PR)和肝脏组织特异性去唾液酸糖蛋白受体(ASGPR)。与在细胞中普遍表达的CI-M6PR相比,ASGPR是一种肝脏组织特异性的LTR,因此基于ASGPR的LYTAC技术是有效且具有特异性的。但是,在疾病模型或临床研究中,靶向ASGPR的LYTAC治疗指数和疗效如何,以及在不同的治疗应用中是否应该选择不同的LTR,仍是一个值得讨论的问题。此外,对LYTAC介导的蛋白质降解过程需要进行更加深入的研究,厘清不同内吞受体在内吞、解离、溶酶体运输以及降解过程中,动力学上是否存在差异,以及这些因素对LYTAC效率的影响。另外,还需要进一步确定LYTAC的靶标以及具体的应用范围。尽管仍有大量亟待解决的问题,但是这些新的发现标志着LYTAC取得了突破性进展,为未来的临床研究开辟了一条更加清晰的道路[15]。
2. 自噬靶向嵌合体(autophagytargeting chimera, AUTAC)技术
自噬靶向嵌合体(AUTAC)是第一个利用溶酶体自噬途径降解目标蛋白的技术。这一概念最初由Takahashi等人[16]于2019年提出。由于内源性核苷酸8-硝基环单磷酸鸟苷(8-nitroguanosine 3′,5′-cyclic monophosphate, 8-nitro-cGMP)是细胞募集自噬体的重要信号分子,在自噬降解中扮演了重要角色,于是Takahashi等人设计了基于内源性修饰cGMP的AUTAC小分子:将靶向蛋白或细胞器的配体与用于募集自噬体(鸟嘌呤衍生物)的标签以linker相连。AUTAC的作用机制是通过降解标签模拟S-鸟苷酸化修饰,诱导靶蛋白的K63多聚泛素化,进而招募自噬体实现溶酶体途径降解。

【图7】AUTAC通过自噬途径选择性降解目标蛋白16]
通过结构优化,作者发现4-氟苄基鸟嘌呤是最优的自噬体募集配体。在此研究基础上设计的系列AUTAC分子(AUTAC1、AUTAC2、AUTAC3)已成功用于降解甲硫氨酸氨基肽酶(methionine aminopeptidase 2, MetAP2)、FK506结合蛋白(506-binding protein, FKB12)、BET家族蛋白等多种疾病相关的靶蛋白的降解。此外,靶向线粒体外膜转运蛋白配体2-苯基-3-乙酮酰胺的AUTAC4(mito-AUTAC)能够实现碎片化线粒体的降解,可以显著提高唐氏综合征成纤维细胞的线粒体活性,显示出AUTAC治疗相关疾病的巨大潜力。

【图8】AUTAC1、AUTAC2、AUTAC3、AUTAC4分子结构[16]

【图9】AUTAC4分子结构及AUTAC4介导线粒体的溶酶体降解 [17]
最近,Pei等人[18]报道了一种新型AUTAC分子,通过靶向自噬关键蛋白LC3诱导BRD4蛋白的自噬降解。他们通过结构改造、活性测试,筛选得到最优化合物10f,在多种肿瘤细胞内(A549, HL60, MDA-MB-231, MCF-7, MDA-MB-436, MDA-MB-468)均能高效降解BRD4蛋白,有效诱导细胞凋亡并延迟G1期,表现出广泛的抗增殖活性。

【图10】靶向LC3的新型AUTAC分子10f [25]
以上结果证明了AUTACs的可行性以及在药物发现方面的巨大潜力,为基于溶酶体自噬途径的蛋白质降解相关药物的开发奠定了基础。但是,未来仍需进一步明确AUTAC的降解潜能(比如对蛋白聚集体的降解作用)、AUTAC具体的降解机制(其多聚泛素化降解标签如何发挥作用)、AUTAC降解的动力学影响因素,为AUTAC的药物开发、临床研究奠定基础[17]。
3. 自噬体偶联化合物(autophagosome-tethering compound, ATTEC)技术
自噬体偶联化合物(ATTEC)同样利用溶酶体自噬途径降解目标蛋白,与AUTAC不同,ATTEC不参与诱导靶蛋白的泛素化,而是直接将靶蛋白和自噬关键蛋白LC3相结合,促进靶蛋白被自噬体吞噬,选择性实现靶蛋白的降解。
2019年,Li等人[19]报道了首例ATTEC系列分子的相关研究。首先,他们提出了如下假设:能够与致病蛋白和自噬关键蛋白LC3相互作用的化合物可能可以将前者结合在吞噬细胞上,以进行后续的自噬降解。接着,他们以突变型亨廷顿蛋白(the mutant HTT, mHTT)为例,验证假设的可行性。mHTT是引发亨廷顿病(Huntington disease, HD)的主要原因,该蛋白具有重复扩展的多聚谷氨酰胺(polyglutamine, polyQ)[20]。作者通过基于小分子微阵列(small-molecule-microarray-based screening, SMMs)和斜入射反射率差异(oblique-incidence reflectivity difference, OI-RD)的高通量筛选,得到了4个能够偶联LC3和mHTT但不与野生型HTT蛋白发生相互作用的化合物。

【图11】ATTEC的发现过程及其作用机制[7]
研究表明,自噬体偶联化合物可以在细胞或者动物模型体内降解mHTT,并减弱HD相关的表型。体外实验表明,ATTEC分子可以特异性识别polyQ并与其发生相互作用,从而选择性降解mHTT,不影响野生型HTT蛋白水平。此外,ATTEC分子还能够降解其他致病的polyQ蛋白,例如会导致小脑性共济失调III型(spinocerebellar ataxia type III)的突变蛋白ATXN3[19]。同时,作者发现ATTEC分子(5和8)可以通过血脑屏障,并可以在约100 nM的浓度下发挥靶向降解蛋白的作用,表明基于ATTEC技术的药物开发存在巨大潜力[21]。
由于ATTEC能够直接与自噬体蛋白LC3相互作用且不经过泛素化过程,因此ATTEC分子在降解DNA/RNA分子、受损的细胞器等需通过自噬识别的非蛋白物质方面具有巨大潜力;ATTEC分子量较低,透膜性好,能够更好地满足药代动力学等方面的要求,可能具有更好的药物特性。但是,目前尚不清楚 ATTEC 分子是否会因其诱导的自噬增加而损害大脑中正常的蛋白质,以及是否存在脱靶效应,对神经系统造成继发性损伤,仍需进行进一步研究。
4. 分子伴侣介导的自噬(chaperone-mediated autophagy, CMA)嵌合体技术
分子伴侣介导的自噬(CMA)是第一个可以用来佐证溶酶体能够选择性降解细胞内成分的例子[22]。CMA介导的蛋白质降解机制如下:首先,分子伴侣热休克蛋白70(heat shock cognate 70 kDa protein, HSC70)识别细胞质内具有KFERQ基序的可溶性蛋白并形成复合物;而后,该复合物与溶酶体膜上的溶酶体相关膜蛋白2A(lysosome-associated membrane protein 2A, LAMP2A)结合,导致LAMP2A低聚化,介导分子伴侣-底物复合物转运至溶酶体中,从而完成底物蛋白的降解[23]。

【图12】CMA的作用机制[22]
21世纪初,Nagai等人[24], [25]发现了一条由46个氨基酸组成、其中含有两个不同HSC70结合基序和两个相同多聚谷氨酰胺结合肽1序列(the polyglutamine-binding peptide 1 sequence, QBP1)的多肽链,能够特异性识别并结合mHTT中的polyQ。在小鼠神经母细胞瘤细胞Neuro2a实验中,该多肽能够通过CMA特异性降解带有polyQ片段的mHTT蛋白。体内研究进一步表明,多肽分子可以改善HD小鼠的病理和表型。为了解释这种现象,作者提出:一种具有HSC70结合基序的类肽分子与目标蛋白的一段特定结构域融合,可能可以治疗由突变蛋白引起的疾病。
2014年,Fan等人[26]基于CMA途径设计了一种嵌合多肽,用于诱导分子伴侣介导的溶酶体自噬。该课题组设计了一个由三个结构域组成的靶向肽:可以跨越血脑屏障和质膜传递的细胞膜穿透结构域(cell membrane-penetrating domain, CMPD)、能够与内源性目标蛋白特异性结合的蛋白质结合域(protein-binding domain, PBD)、CMA靶向基序(CMA-targeting motif, CTM)。作者假设靶向肽将通过CMPD进入细胞,经由PBD特异性识别与目标蛋白形成稳定的复合物,通过CTM-CMA的作用将肽-蛋白质复合物转运至溶酶体从而完成降解。该设计除了实现死亡相关蛋白激酶1(DAPK1, 160 kDa)的靶向降解外,还成功应用到α-突触核蛋白(α-synuclein, 18 kDa)和骨架蛋白PSD-95(95 kDa)的靶向降解。
2018年,Wang等人[27]发现了PD-L1的溶酶体降解途径及其调节因子HIP1R。HIP1R上的多肽基序MDFSGLSLIKLKKQ具有与KFERQ基序类似的溶酶体靶向活性,可应用于溶酶体靶向的蛋白质降解。研究发现,HIP1R可以直接与PD-L1发生作用,并通过溶酶体靶向信号通路将其转运至溶酶体发生降解,以增强T淋巴细胞介导的细胞毒性,发挥抗肿瘤作用。由于HIP1R的功能依赖于两个序列延伸——一个与PD-L1发生特异性识别,另一个介导PD-L1向溶酶体运输——于是作者设计了PD-LYSO多肽,可以靶向肿瘤细胞中的PD-L1转运至溶酶体发生降解。HIP1R介导的PD-L1溶酶体降解的发现,为抑制PD-L1作为免疫治疗的关键靶点提供了一条潜在的新途径。

【图13】HIP1R介导PD-L1发生溶酶体降解 [27]
2019年,Zhou等人[28]设计了由穿膜肽Tat、细胞周期蛋白依赖性激酶5(cyclin-dependent kinase 5, CDK5)的配体和CMA靶向基序CTM组成的Tat-CDK5-CTM嵌合多肽,该多肽能够与CDK5特异性结合并介导其降解,阻断了CDK5与小鼠NMDA受体B亚单位NR2(NR2B)的相互作用,减少了小鼠脑部神经元的死亡并对其神经功能有一定程度的改善。这是一种具有优良前景的中风治疗干预措施,并且,通过CMA途径实现对CDK5蛋白的成功降解,提供了一种调节内源蛋白水平的新方法。

【图14】Tat-CDK5-CTM破坏NR2B和CDK5的结合并导致CDK5降解[28]
CMA嵌合多肽的设计,为蛋白质错误折叠引起的相关疾病的治疗提供了新的策略,促进了潜在的药物开发。靶向CMA的嵌合体技术具有更快的降解速度、更优的可逆性和剂量依赖性、更强的专一性。然而,该技术目前也存在着一些局限性和挑战,仍有待进一步改进:(1)设计的嵌合多肽与靶蛋白应具有高亲和力的结构域;(2)嵌合多肽存在的跨膜能力差和稳定性低等问题是影响其最终成药的主要障碍
结 语
相较于目前炙手可热的PROTAC技术,靶向溶酶体降解技术具有更加广阔的底物适用范围,它不仅可以降解胞质蛋白,而且可以降解胞外蛋白、蛋白聚集体、DNA/RNA分子、过氧化物酶体、核糖体、受损的线粒体或微生物病原体等等,这是蛋白酶体途径降解技术不能完成的目标。
目前,已经产生并发展的主要有溶酶体靶向嵌合体(lysosome-targeting chimera, LYTAC)技术、自噬靶向嵌合体(autophagytargeting chimera, AUTAC)技术、自噬体偶联化合物(autophagosome-tethering compound, ATTEC)技术以及分子伴侣介导的自噬(chaperone-mediated autophagy, CMA)嵌合体技术,用于降解细胞外蛋白、大分子或受损的细胞器。
上述四种方法各有优势,同样均存在局限性和挑战:LYTAC能够降解胞外蛋白和膜蛋白,但是其分子量大、降解速率慢,安全性、选择性、药代动力学特性均需要得到进一步改善;AUTAC具备降解受损的线粒体等细胞器的能力,但是由于目前作用机制不清晰导致研究受到限制;ATTEC分子量小,能够较好地透过血脑屏障,在降解神经系统疾病中的靶蛋白、编码致病蛋白的遗传物质等方面有着巨大潜力,但是目前尚不清楚 ATTEC 分子是否会因其诱导的自噬增加而损害大脑中正常的蛋白质,以及是否存在脱靶效应;CMA降解速度快、专一性强,在治疗因蛋白质错误折叠引起的相关疾病(如阿尔茨海默病[29])方面有着巨大的潜力,但透膜性差、稳定性低,需要进一步改善以实现相关药物的开发。
综上,虽然靶向溶酶体降解技术的相关研究目前仍处于早期阶段,但是我们相信,随着科研工作者们不断的探索,克服种种局限与挑战,靶向溶酶体降解技术将为先前人们无法战胜的疾病提供新的治疗策略,与之相关的药物发现也将出现新的热潮。
参考资料
[1] Tang, M., et al., Proteasome functional insufficiency activates the calcineurin-NFAT pathway in cardiomyocytes and promotes maladaptive remodelling of stressed mouse hearts. Cardiovasc Res, 2010. 88(3): p. 424-33.
[2] Dong, G., et al., Molecular Glues for Targeted Protein Degradation: From Serendipity to Rational Discovery. J Med Chem, 2021. 64(15): p. 10606-10620.
[3] Tooze, S.A., A. Abada, and Z. Elazar, Endocytosis and autophagy: exploitation or cooperation? Cold Spring Harb Perspect Biol, 2014. 6(5): p. a018358.
[4] di Ronza, A., et al., CLN8 is an endoplasmic reticulum cargo receptor that regulates lysosome biogenesis. Nature Cell Biology, 2018. 20(12): p. 1370-+.
[5] Ni, X., M. Canuel, and C.R. Morales, The sorting and trafficking of lysosomal proteins. Histology and Histopathology, 2006. 21(8): p. 899-913.
[6] Malini, E., et al., Role of LIMP-2 in the intracellular trafficking of beta-glucosidase in different human cellular models. Faseb Journal, 2015. 29(9): p. 3839-3852.
[7] Pei, J.P., et al., Targeting Lysosomal Degradation Pathways: New Strategies and Techniques for Drug Discovery. Journal of Medicinal Chemistry, 2021. 64(7): p. 3493-3507.
[8] Xiang, H.G., et al., Targeting autophagy-related protein kinases for potential therapeutic purpose. Acta Pharmaceutica Sinica B, 2020. 10(4): p. 569-581.
[9] Stolz, A., A. Ernst, and I. Dikic, Cargo recognition and trafficking in selective autophagy. Nature Cell Biology, 2014. 16(6): p. 495-501.
[10] Dikic, I. and Z. Elazar, Mechanism and medical implications of mammalian autophagy. Nature Reviews Molecular Cell Biology, 2018. 19(6): p. 349-364.
[11] Banik, S.M., et al., Lysosome-targeting chimaeras for degradation of extracellular proteins. Nature, 2020. 584(7820): p. 291-297.
[12] Ahn, G., et al., LYTACs that engage the asialoglycoprotein receptor for targeted protein degradation. Nature Chemical Biology, 2021. 17(9): p. 937-+.
[13] Zhou, Y., et al., Development of Triantennary N-Acetylgalactosamine Conjugates as Degraders for Extracellular Proteins. ACS Cent Sci, 2021. 7(3): p. 499-506.
[14] Caianiello, D.F., et al., Bifunctional small molecules that mediate the degradation of extracellular proteins.Nature Chemical Biology, 2021. 17(9): p. 947-+.
[15] Paulk, J., PROTEIN DEGRADATION Lysosome-targeting chimeras evolve. Nature Chemical Biology, 2021. 17(9): p. 931-933.
[16] Takahashi, D., et al., AUTACs: Cargo-Specific Degraders Using Selective Autophagy. Molecular Cell, 2019. 76(5): p. 797-+.
[17] Takahashi, D. and H. Arimoto, Targeting selective autophagy by AUTAC degraders. Autophagy, 2020. 16(4): p. 765-766.
[18] Pei, J., et al., Developing potent LC3-targeting AUTAC tools for protein degradation with selective autophagy.Chem Commun (Camb), 2021. 57(97): p. 13194-13197.
[19] Li, Z., et al., Allele-selective lowering of mutant HTT protein by HTT-LC3 linker compounds. Nature, 2019. 575(7781): p. 203-209.
[20] Li, Z., et al., ATTEC: a potential new approach to target proteinopathies. Autophagy, 2020. 16(1): p. 185-187.
[21] Ding, Y., Y. Fei, and B. Lu, Emerging New Concepts of Degrader Technologies. Trends Pharmacol Sci, 2020. 41(7): p. 464-474.
[22] Kaushik, S. and A.M. Cuervo, The coming of age of chaperone-mediated autophagy. Nat Rev Mol Cell Biol, 2018. 19(6): p. 365-381.
[23] Li, W., et al., Chaperone-mediated autophagy: Advances from bench to bedside. Neurobiol Dis, 2019. 122: p. 41-48.
[24] Nagai, Y., et al., Inhibition of polyglutamine protein aggregation and cell death by novel peptides identified by phage display screening. J Biol Chem, 2000. 275(14): p. 10437-42.
[25] Nagai, Y., et al., Prevention of polyglutamine oligomerization and neurodegeneration by the peptide inhibitor QBP1 in Drosophila. Hum Mol Genet, 2003. 12(11): p. 1253-9.
[26] Fan, X., et al., Rapid and reversible knockdown of endogenous proteins by peptide-directed lysosomal degradation.Nat Neurosci, 2014. 17(3): p. 471-80.
[27] Wang, H., et al., HIP1R targets PD-L1 to lysosomal degradation to alter T cell-mediated cytotoxicity. Nat Chem Biol, 2019. 15(1): p. 42-50.
[28] Zhou, Y.F., et al., The Peptide-Directed Lysosomal Degradation of CDK5 Exerts Therapeutic Effects against Stroke.Aging Dis, 2019. 10(5): p. 1140-1145.
[29] Bourdenx, M., et al., Chaperone-mediated autophagy prevents collapse of the neuronal metastable proteome. Cell, 2021. 184(10): p. 2696-2714 e25.
不感兴趣
看过了
取消

 打赏
打赏
不感兴趣
看过了
取消
精彩评论
相关阅读




中国医院排行榜















 010-82736610
010-82736610
 股票代码: 872612
股票代码: 872612



©2012-2023 北京华媒康讯信息技术股份有限公司 All Rights Reserved. 注册地址:北京 联系电话:82736610
 京公网安备 11010802020745号
京公网安备 11010802020745号


